It is through our study of our hominin ancestors and relatives that we are exposed to a world of “might have beens”: of other paths not taken by our species, other ways of being human. But in order to better understand these different evolutionary trajectories, we must first define the terms we are using. If an imaginary line were drawn between ourselves and our closest relatives, the great apes, bipedalism (or habitually walking upright on two feet) is where that line would be. Hominin, then, means everyone on “our” side of the line: humans and all of our extinct bipedal ancestors and relatives since our divergence from the with chimpanzees.
Historic interpretations of our evolution, prior to our finding of early hominin fossils, varied. Debates in the mid-1800s regarding hominin origins focused on two key issues:
Where did we evolve?
Which traits evolved first?
Charles Darwin hypothesized that we evolved in Africa, as he was convinced that we shared greater commonality with chimpanzees and gorillas on the continent (Darwin 1871). Others, such as Ernst Haekel and Eugene Dubois, insisted that we were closer in affinity to orangutans and that we evolved in Eurasia where, until the discovery of the Taung Child in South Africa in 1924, all humanlike fossils (of Neanderthals and Homo erectus) had been found (Shipman 2002). Adding to this debate was the discovery of the Piltdown Man in England, which turned out later to be a forgery of a modified orangutan mandible and medieval human skull.
Within this conversation, naturalists and early paleoanthropologists (people who study human evolution) speculated as to which human traits came first. These included the evolution of a big brain (encephalization), the evolution of the strange way in which we move about on two legs (bipedalism), and the evolution of our strange flat faces and small teeth (indications of dietary change). Original hypotheses suggested that in order to be motivated to change diet and move about in a bipedal fashion, the large brain needed to have evolved first. And, until research picked up in Africa, fossil finds of species mentioned above predominantly had larger heads.
We now know that bipedal locomotion is one of the first things that evolved in our lineage, with early relatives having small brains and more apelike dentition. In this chapter, we will tease out the details of what this looks like in terms of morphology (i.e. the study of the form or size and shape of things; in this case, skeletal parts).
We also know that early human evolution occurred in a very complicated fashion. We have multiple species (multiple genera), diverse in the extent to which they move like us and the diets on which they subsist. Specimen finds have been made all along the East African Rift System (EARS; in Ethiopia, Kenya, Tanzania, and Malawi), in limestone caves in South Africa, and in Chad. Dates of these early relatives range from around 7 million years ago (mya) to around 1 mya, overlapping temporally with members of our genus, Homo (Figure 9.1).
Figure \(\PageIndex{1}\): East Africa Rift System.
Yet there is still so much to understand. Modern debates now look at the relatedness of these species to us and to one another. Discussions regarding which of these species were able to make and use tools continue. Every site discovery in the patchy hominin fossil record tells us more about our evolution. New scientific techniques provide us with insight into the diets, environments, and lifestyles of these ancient relatives that was not available to researchers even ten years ago.
A Note on Brain Size
It is worth noting that while brain size expansion is seen primarily in our genus, Homo, earlier hominin brain sizes were highly variable between and within taxa, from 300 cc (cranial capacity, cm3), estimated in Ardipithecus, to 550 cc, estimated in Paranthropus boisei. The lower estimates are well within the range of variation of nonhuman extant great apes, and body size variability also plays a role in the interpretation of whether brain size could be considered large or small for a particular species or specimen.
Increases in brain size do not necessarily correlate with an increase in intelligence in animals, especially if body size is not taken into consideration. However, the brain is an expensive tissue to build and maintain. Researchers therefore argue that the cost of maintenance must yield some evolutionary benefit. This is more easily understood in hominins where the stone tool record (an indication of behavior and intelligence) is well associated with the species.
Ancestral and Derived Traits
In Chapter 5, you were introduced to ways of organizing living taxa. In the past, taxonomy was primarily based on morphology (i.e., the physical features of organisms). Today they are tied to known relationships based on molecular phylogeny (such as based on DNA) or a combination of the two. This technique is complicated when applied to living taxa, but it becomes immensely more difficult when we seek to categorize ancestor-descendant relationships in long-extinct forms, where molecular information is no longer preserved. In many ways, we find ourselves falling back on morphological comparisons (often on teeth and partially fossilized skeletal material) in the absence of genetic material.
It is here that we turn to the related concepts of cladistics and phylogenetics. Cladistics groups organisms according to their last common ancestors based on shared derived traits. These are traits (in the case of early hominins, morphological) that are evolved, differing from those seen in earlier populations or forms. These new or modified traits or characteristics provide evidence of evolutionary relationships, and organisms with the same derived traits are grouped in the same clade (Figure 9.2). For example, if we use feathers as a trait, we can group pigeons and ostriches into the clade “birds.” A good example we will see in this chapter is the grouping of what is known as the “Robust Australopithecines,” whose cranial and dental features differ from those of other hominins of a similar or earlier time period and can therefore be considered derived.
Figure \(\PageIndex{2}\): Clades refer to groups of species or taxa that share a common ancestor.
It is also worth noting that species designations for early hominin specimens are often highly contested. This is, in part, due to the fragmentary nature of the fossil record, the large timescale (in millions of years) with which paleoanthropologists need to work, and because of the difficulty in evaluating whether morphological differences and similarities are due to meaningful phylogenetic or biological differences or subtle differences/variation in niche occupation or time. In other words, do morphological differences indicate different species? How would classifying species in the paleoanthropological record compare with classifying living species today, for whom we can sequence genomes and observe lifestyles?
There are also broader philosophical differences among researchers when it comes to paleo-species designations. Some scientists, known as “lumpers,” argue that large variability is expected among multiple populations in a given species over time. These researchers will therefore prefer to “lump” specimens of subtle differences into single taxa. Others, known as “splitters,” argue that species variability can be measured and that even subtle differences can imply differences in niche occupation that are extreme enough to mirror modern species differences. In general, splitters would consider geographic differences among populations as meaning that a species is polytypic (i.e., capable of interacting and breeding biologically but having morphological population differences). This is worth keeping in mind when learning about why species designations may be contested.
This further plays a role in evaluating ancestry. Debates over which species “gave rise” to which continue to this day. It is common to try to create “lineages” of species—determining, in other words, when one species evolves into another over time. We refer to these as chronospecies. Constructed hominin phylogenetic trees are routinely variable, changing with new specimen discoveries, new techniques for evaluating and comparing species, and, some have argued, nationalist or biased interpretations of the record. More recently, some researchers have shifted away from “treelike” models of ancestry toward more nuanced metaphors such as the “braided stream,” where some levels of interbreeding among species and populations are seen as natural processes of evolution.
Finally, it is worth considering the process of fossil discovery and publication. Some fossils are easily diagnostic to a species level and allow for easy and accurate interpretation. Some, however, are more controversial. This could be because they do not easily preserve or are incomplete, making it difficult to compare and place within a specific species (e.g., the patella or knee bone). Researchers often need to make several important claims when announcing or publishing a find: a secure date (if possible), clear association with other finds, and an adequate comparison among multiple species (both extant and fossil). It is therefore not uncommon for the scientific community to know that an important find was made years before it is scientifically published.

 Figure \(\PageIndex{1}\): East Africa Rift System.
Figure \(\PageIndex{1}\): East Africa Rift System.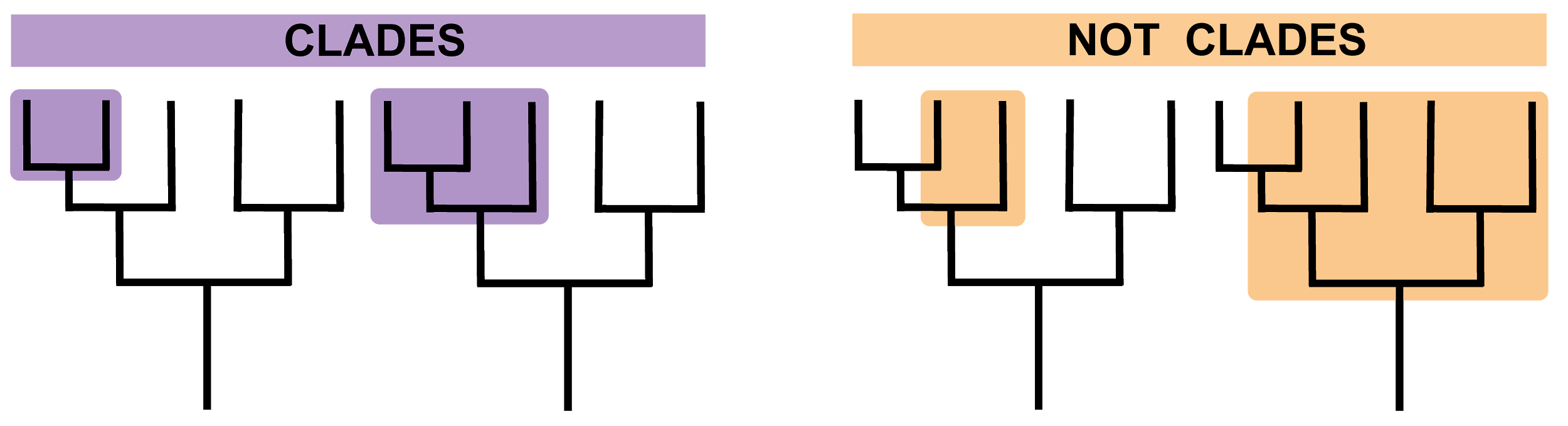 Figure \(\PageIndex{2}\): Clades refer to groups of species or taxa that share a common ancestor.
Figure \(\PageIndex{2}\): Clades refer to groups of species or taxa that share a common ancestor.