Reconstruction of Homo erectus. “Homo erectus new” by Lillyundfreya is licensed under CC BY-SA 3.0.
SITES
Algeria: Tighenif
Morocco: Thomas Quarries and Sidi Abderrahman
Ethiopia: Konso Gardula and Omo
Kenya: Olorgesaillie and Nariokotome
Tanzania: Olduvai Gorge
South Africa: possibly Swartkrans
PEOPLE
Richard Leakey, Kimoya Kimeu, Bernard Ngeneo
As mentioned, the African form of the “erectus grade” is termed Homo ergaster.
PHYLOGENY
H. ergaster is thought to have evolved from either H. habilis or H. rudolfensis in East Africa. However, it is possible that H. habilis may have been the first to leave Africa, after which it may have evolved into a pre-ergaster/erectus form that then moved into Africa and Asia. If H. habilis was in our ancestry, the latter scenario might explain how the more modernly proportioned H. ergaster appeared in the fossil record contemporary with H. habilis in East Africa. Of course, the hit and miss nature of the fossil record and fossil and species discoveries could also explain that phenomenon, i.e. the seeming lack of transitional forms in Africa.
DISCOVERY AND GEOGRAPHIC RANGE
The earliest H. ergaster material is from the East Lake Turkana site of Koobi Fora in Kenya. Richard Leakey is credited with this 1.8 mya discovery. Other sites outside of Africa are contemporary with African sites, e.g. the 1.8 mya Dmanisi site in the Republic of Georgia and the 1.8–1.6 mya site of Modjokerto in Java. (Note: There are problems with the Javanese dates because the fossil-containing layers are not conducive to more reliable dating methods.) The almost complete Nariokotome or Turkana Boy (see Figure 28.2) from the West Lake Turkana region of Kenya was discovered in 1984 by Kamoya Kimeu and dated to 1.6 mya. The skeleton has been extremely important for reconstructing body morphology and limb proportions. The boy is thought to have been eight years old based upon tooth development patterns. He was formerly thought to be as old as 15, based on his height, stage of bone development, and hypothesized growth trajectories. However, dental calculations can accurately determine age due to the daily pattern of enamel deposition during tooth development. Scientists can count the microscopic, bead-like deposits that are laid down daily during the course of a tooth’s development. Once it was determined that he was only eight years old yet 5´3˝ tall, it was apparent that H. ergaster developed at a much faster rate, more like a chimp than a human. Had Turkana Boy lived to adulthood, he would have been over 6´ tall. His morphology was adapted to the hot, dry conditions in equatorial East Africa, i.e. tall and long-limbed, similar to modern peoples of the region.
Other African sites include the North African sites of Tighenif (formerly Ternifine and sometimes assigned to Homo mauritanicus) in Algeria and Thomas Quarries and Sidi Abderrahman in Morocco; the East African sites of Konso Gardula and Omo in Ethiopia, Olorgesaillie in Kenya, and Olduvai Gorge in Tanzania; and possibly the South African site of Swartkrans, although it is not universally accepted that H. ergaster was there.
“Turkana Boy” by Mike Peel is licensed under CC BY-SA 4.0.
PHYSICAL CHARACTERISTICS
H. ergaster exhibited robust craniofacial characteristics relative to modern humans, but overall we see the continued reduction in dentition and masticatory apparatus. Their brains were large, with a maximum cranial capacity of 1200 cc and a range of 800–1200 cc, based on the material from the three continents. However, the relative brain size of early specimens may not have been much greater than that seen in early Homo. They had fairly robust supraorbital and nuchal regions. The skull vault was long and low, termed platycephalic, with a low maximum width. Our vaults are much higher, and we have high maximum width, due to subsequent cerebral expansion. The shape of the vault in cross-section has been described as that of a turtle shell. H. ergaster, like all hominins before and especially the robust australopiths, exhibits postorbital constriction between the orbits and the cranial vault. Their temporal lines were more pronounced than ours (see Figure 28.4), indicating that they had more powerful jaw muscles and chewing capabilities. They did not have much of a forehead due to several of the aforementioned craniofacial characteristics. Their nasal bones suggest a human-like, projecting nose. This would have been adaptive in arid as well as cold conditions. In addition to warming and humidifying inhaled air, the moisture in warm exhaled air condenses on the cooler nasal membranes, resulting in water conservation. The jaws were somewhat prognathic in the alveolar region, i.e. the bone that houses the teeth, and the mandibles were robust. They lacked a chin, a characteristic seen only in anatomically modern humans.
Relative to H. habilis, long thought to be the species from which H. ergaster was derived, H. ergaster possessed a longer skull vault, a larger brain (however, see previous paragraph), a smaller temporal fossa (the opening formed by the zygomatic arch where jaw muscles pass through and/or attach), a shorter face, a larger nose, reduced dentition and jaw robusticity, and a heavier nuchal region.
Relative to Asian H. erectus, H. ergaster possessed a higher cranial vault, and the bones of the vault were thinner. They also did not exhibit the sagittal keel (see Homo erectus, Chapter 29) typical of the Asian form. However, the Koobi Fora specimen exhibits slight keeling (see Figure 28.4).
Koobi Fora Homo ergaster. Postorbital constriction. Slight keeling Temporal line. “Homo ergaster skull replica, World Museum Liverpool” by Reptonix is licensed under CC BY 3.0.
Postcranially, H. ergaster was very human-like. While the thorax may still have been somewhat conical, they had more of a waist, demonstrating an uncoupling of the lower limb from the torso. The small birth canal relative to adult brain size suggests a long period of postnatal brain growth, as seen in subsequent hominins, especially our own species. They were tall, achieving heights of over 6´. They were also more robust than premodern humans that, in turn, were more robust than modern humans. Males were 20–30% larger than females. While the lifespan tended to be short, some individuals lived to be 50 to 60 years of age. There was high infant mortality, with 40% of fossil remains estimated to be less than 14 years of age.
Note
The ideal gas law is easy to remember and apply in solving problems, as long as you get the proper values a
Review of Primitive Characteristics
Retain:
Initially brain possibly no larger than early Homo.
Some robusticity in cranium and face.
Supraorbital and nuchal tori.
Postorbital constriction.
Low forehead.
Conical thorax.
Relative to later hominins:
Platycephalic skull vault with low maximum breadth.
Pronounced temporal lines.
Alveolar prognathism.
Large teeth but smaller than previous species.
No chin.
Review of Derived Characteristics
Protruding nose.
Postcranially robust.
Tall with long legs.
Development of waist (decoupling between torso and lower limb).
ENVIRONMENT AND WAY OF LIFE
The environment of East Africa during the Pleistocene was hot and arid. Savannas had expanded and forests had become increasingly fragmented. There was increased volcanic activity in the Great Rift zone, and that is one hypothesis as to why at least a portion of the population moved out of the area. It is of great interest as to how and why H. ergaster (or a predecessor) first left East Africa. The “how” is not as difficult as the “why.” While they may have crossed the Red Sea, they could also have come up through the Nile Valley and exited Africa across the Arabian land bridge that was exposed at the time. As to why they left, there are a variety of ideas that are not mutually exclusive. They may have ventured farther and farther due to (1) competition for resources with their own or other species of hominins or non-hominins; (2) local resource depletion; (3) similar environment (there is speculation that they were a grassland species that followed African grasslands into West Asian steppes, through the Indian subcontinent, and southeast to Indonesia); (4) following herd animals north and out of Africa; and/or (5) the aforementioned volcanic activity.
While there is debate as to whether the various “erectus” species hunted large game, it is not likely, since they lacked weaponry. It is generally accepted that they were mobile scavengers and foragers that hunted opportunistically. They certainly would have been capable of taking small prey and older and/or injured animals. Faunal remains at the Zhoukoudian, China, site consisted of deer, elephant, rhino, and beaver remains, but the bulk of the animal matter in their diet is thought to have come from smaller animals, such as rodents, hedgehogs, hares, and frogs, as well as ostrich eggs. However, Bunn and Gurtov (2014) have analyzed faunal remains at the Zinj FLK site in Olduvai Gorge, Tanzania (1.8 mya) and concluded that the high percentage (72%) of prime-aged large prey (especially bovids with weights of 250–750 lb) conforms solely to hominin hunting patterns, versus accidental deaths or carnivore kills. Thus at least some H. ergaster are thought to have been practicing ambush hunting and were therefore big-game hunters.
Acheulian hand axe. “Bifaz en mano” by José-Manuel Benito Álvarez is licensed under CC BY-SA 2.5.
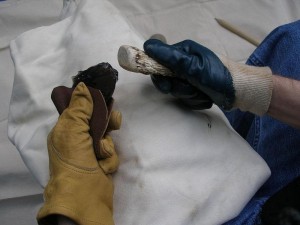
While nomadic, they are thought to have stayed in an area for at least short periods of time, relative to past species. Early H. ergaster is associated with the Oldowan technology, and that is the technology that they took with them out of Africa. H. ergaster subsequently invented a tool tradition, termed Acheulian, that first appears in the archaeological record at 1.4 mya (newer data suggests possibly as early as 1.7 mya) and lasted to as late as 115 kya in some areas. The latter industry spread throughout Africa and as far east as the Indian subcontinent and west to Western Europe. It involved the use of better stone resources and tools that were more refined and standardized than in the Oldowan tradition. The most representative tool was a bifacially worked (shaped on both sides) hand axe in the shape of a teardrop (see Figure 28.5). The tool manifests what is called a “target design,” in that the manufacturer knew what he or she was making and followed standardized steps in shaping the core into the resulting tool form. Once the core was prepared with the use of a stone hammer, the next step involved a “soft hammer” technique (see Figure 28.6). This required the use of a bone or antler hammer that could more carefully control the size and shape of the flakes to be removed on either side of the axe as it was shaped. Sharper and straighter edges resulted from the technique. The hominins had different tools for different purposes. The categories of tools included hand axes, smaller flake tools, choppers, picks, cleavers, and some bone tools. They would likely have been used to kill; butcher; scrape and cut hides; shape wood, bone, antler, and stone; and harvest and process plant foods.
Soft hammer percussion. “Soft Hammer” by ZenTrowel is in the public domain.
It would have been necessary for H. erectus to have had some sort of body covering prior to moving north into colder regions of Eurasia. There is some evidence to support H. erectus’s use of fire, but it is scant. However, even in later species wherein we know they had fire, such as H. heidelbergensis and H. neanderthalensis, there are fewer hearths apparent at archaeological sites than would be expected.
It was previously believed that the erectus grade lacked the technology to cross open water and therefore were limited to land travel. However, the discovery of a site dated to >8 kya on the island of Flores suggests that early H. erectus were capable of crossing fairly sizeable stretches of open sea. It is thus possible that H. ergaster may have rafted across the Red Sea to the Arabian Peninsula or across the Mediterranean to Gibraltar or Italy (via Sicily) during periods of lower sea levels when much of the earth’s water was tied up in ice.
While it is likely that H. ergaster and related species had some complex form of communication, due to their advanced cultural achievements, it is not known whether they had speech or language similar to our own. They possessed the arched palate, flexed cranial base, and cerebral centers that are necessary for speech production and comprehension. However, Turkana Boy had small intervertebral foramina (the openings in between the vertebrae where the spinal nerves exit the spinal cord) in the thoracic region. It is thus thought that they may not have had the necessary motor function for controlled breathing and speech. However as mentioned in the section on Homo habilis (Chapter 23), I believe the standardized nature of the Acheulian tool industry demonstrates active teaching and thus theory of mind that in turn assumes some sort of language.