Explain the basic trends of human evolution including bipedalism and encephalization.
Describe hominins and what distinguishes hominins from other members of the Primate order.
Describe the category of features that distinguishes proto-hominins from hominins.
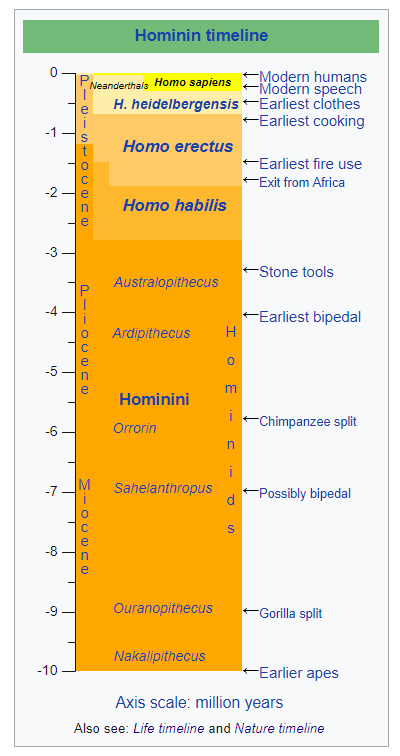
Describe, including approximate date ranges, the evolution of the genus Homo, including early Homo species and modern humans.
Explain how material culture helps inform us about the psychology of ancestral Homo sapiens and other hominin species.
Overview
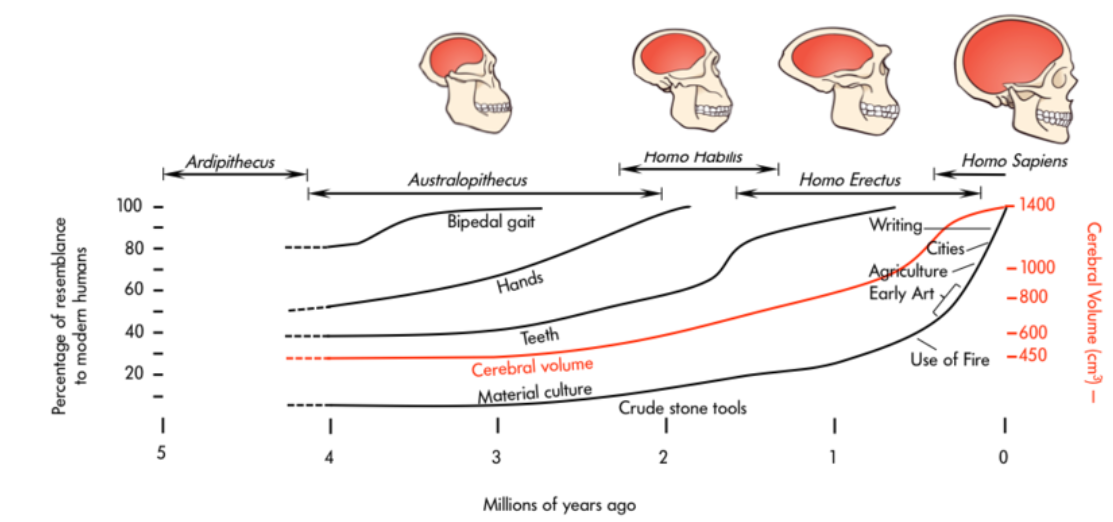
Trends: There are a number of trends in the evolution of the proto-hominins (primitive ape-like species regarded as possibly ancestral to modern humans) to modern Homo sapiens. These traits do not occur all at once, but over millions of years.
Proto-Hominins and Hominins: In determining what fossil features a specimen must have in order to be classified as a hominin (the term used for humans and their ancestors after the split with chimpanzees and bonobos), many different characteristics are examined, including those related to bipedalism and dental features related to chewing. Apes have a space between biting teeth and grinding teeth where the upper canine fits when the jaws close. Other characteristics such as brain and body size are also considered.
Homo Genus: The emergence of the genus Homo (our genus) marks the advent of larger brains, the emergence of material culture (e.g. stone tools), and the eventual colonization of the world outside of Africa.
Material Culture: The earliest evidence of material culture is in the form of stone tools found on sites which dated back to 2.4 million years ago (see Supplementary Content, Chapter 18, Material Culture).
Figure \(\PageIndex{1}\): The original complete skull (without upper teeth and mandible) of a 2.1 million years old Australopithecus africanus specimen so-called "Mrs. Ples" (catalogue number STS 5, Sterkfontein cave, hominid fossil number 5), discovered in South Africa . Collection of the Transvaal Museum, Northern Flagship Institute, Pretoria, South Africa. (CC BY-SA 4.0; José Braga; Didier Descouens).
Human and Ape Evolutionary Paths: Divergence from Common Ancestors
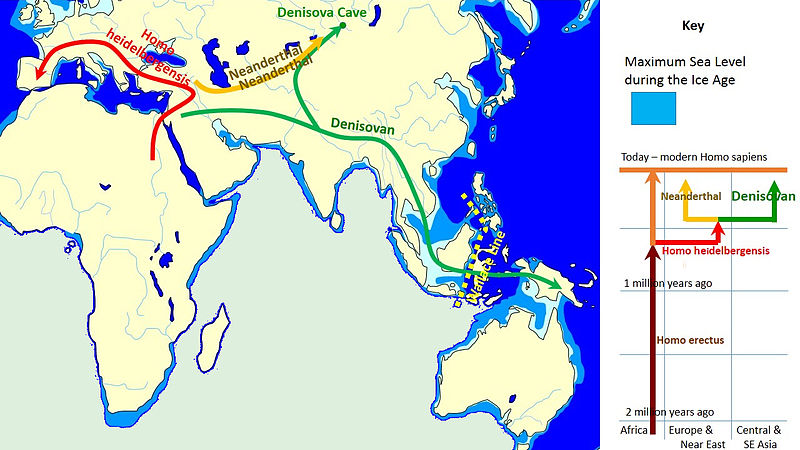
Before we begin our discussion of human evolution, it will be helpful to take a look at the big picture. The two figures below along with Figure 3.7.16, much further below, collectively depict 20 million years of evolution leading to our species, Homo sapiens. Figure 3.7.2 shows the last 10 million years of humanoid evolution. Figure 3.7.3 shows the last 20 million years of humanoid evolution and so is inclusive of Figure 3.7.2, which shows greater detail beginning with the Homininae (African hominids including humans) phylogeny, 10 Mya to present. Both of these figures show our genus Homo at the top left of each diagram. Figure 3.7.16 shows an enlarged and much more detailed depiction of the genus Homo including relationships between Homo sapiens and Neanderthals and the fact that these two species once co-existed at the same time outside of Africa about 30,000-40,000 years ago. In the first two figures immediately below, notice the great divergence into a surprisingly large number of species over millions of years. Also notice how small and how recent the genus Homo is by comparison, and how one very recent species that emerged during the Pleistocene, humans, has so quickly come to dominate the Earth, at least for now. It is hoped that by closely examining these figures, you will be better able to organize the details that follow in this section and that you will be able to understand them within the larger context of primate evolution.
Figure \(\PageIndex{2}\): Hominini (includes Homo and Pan, but excludes gorillas) and Homininae (African hominids including humans) phylogeny, 10 Mya to present; the human branch beginning with Homo, top left, is enlarged and shown in greater detail in Figure 3.7.16 closer to the bottom of this page. The Hominin ancestral line (modern humans, extinct human species and all our immediate ancestors including members of the genera Homo, Australopithecus, Paranthropus and Ardipithecus)and the ape line, including gorillas, diverged from one another about 8 Mya (million years ago) and the Hominin and Pan (chimpanzee) lines diverge from one another about 6 Mya. Homo sapiens are shown at the very top left. To go back 20 Mya, see Figure 3.7.3 below. There you can see that Pongo (orangutans) diverged from the Hominin line much earlier, about 14 million years ago (Image from Wikimedia Commons; File:Hominini lineage.svg; https://commons.wikimedia.org/wiki/F...ni_lineage.svg; by Dbachmann; licensed under the Creative CommonsAttribution-Share Alike 4.0 International license).
Figure \(\PageIndex{3}\): Hominoidea phylogeny, 20 Mya to present. A hominoid, sometimes called an ape, is a member of the superfamily Hominoidea: extant members are the gibbons (lesser apes, family Hylobatidae) and the hominids. A hominid is a member of the family Hominidae, the great apes: orangutans (Ponginae; Pongo), gorillas (Gorillini), chimpanzees (Pan) and humans. Note that the split between the Homo line, leading to humans, from the chimpanzee and gorilla lineages occurred about six million and eight million years ago, respectively. (Image from Wikimedia Commons; File:Hominoidea lineage.svg; https://commons.wikimedia.org/wiki/F...ea_lineage.svg; by Dbachmann; licensed under the Creative CommonsAttribution-Share Alike 4.0 International license).
Trends in Human Evolution
A number of questions about human evolution are:
Why did our earliest ancestors stand up?
Why did some species become extinct until only one species, Homo sapiens, was left?
When, where and why did modern humans evolve?
What was the role of the Neanderthals?
What makes us human?
While hypotheses suggest answers, research continues to refine our understanding of human evolution.
This section provides an overview of key finds and trends in hominin evolution. (For additional information, see the "For Further Exploration" at the end of this page).
Figure \(\PageIndex{4}\): A man contemplating his evolution.
Defining Hominins
It is through our study of our hominin ancestors and relatives that we are exposed to a world of “might have beens”--of other paths not taken by our species, of other ways of being human. But in order to better understand these different evolutionary trajectories, we must first define the terms we are using. If an imaginary line were drawn between ourselves and our closest relatives, the great apes, bipedalism (habitually walking upright on two feet) is where that line would be drawn. Hominin, then, means everyone on “our” side of that line defined by bipedalism. Thus, hominins are humans and all of our extinct bipedal ancestors and relatives since our divergence from the last common ancestor (LCA) that we shared with chimpanzees. As shown above in Figures 3.3.2 and 3.3.3, many scientists believe that this split between human and chimpanzee lineages took place about 6 million years ago. However, data from DNA using estimated mutation rates suggests that the split between the human and ape lineages may have occurred much earlier--divergence between human and chimpanzee lines taking place about 7 to 8 million and possibly as many as 13 million years ago, while the divergence between human and gorilla lines may have occurred as long ago as 8 to 19 million years ago (Gibbons, 2012). These different estimates may be confusing to students. However, this wide range of estimates serves as a good example of how our knowledge is incomplete and sometimes changing as new discoveries and methods arise. In this context, it is best to keep in mind that "[…] when all is said and done a taxonomy is just a hypothesis; it is not written on stone tablets" (Wood, 2010, p. 8908).
Morphological Trends in Human Evolution
There are a number of trends in the evolution of the human lineage from the proto-hominins (early human ancestral species) to modern Homo sapiens. These traits do not occur all at once, but over millions of years. In general, the trends include:
the forward movement of the foramen magnum (the hole at the bottom of the skull where spinal cord enters skull to connect with brain): related to upright posture
a reduction in the size of the canines
an increase in the size of the molars
an increase in cranial capacity
flattening of the face
rounding of the skull
Again, not all of these traits occur at the same time and there is variation among the various hominin species, but all of these morphological characteristics occur in the evolutionary line of Homo sapiens. Two other trends are especially important in the evolution of hominins:
bipedalism, and
encephalization of the brain.
These are discussed in more detail next.
Bipedalism
Bipedalism, or upright walking, was the first morphological trait on the road to humanity. Human bipedalism is the primary form of moving around (this is called habitual bipedalism). Other primates practice temporary or occasional bipedal locomotion, e.g., primates like chimpanzees may walk bipedally while they carry something in their hands. Few other animals are habitual bipeds like humans, e.g., birds and kangaroos.
Figure \(\PageIndex{5}\): Human skeleton walking.
There are numerous anatomical changes that evolved to make hominins efficient bipedal locomotors including changes in position of foramen magnum (see above), the vertebral column of the spine, the pelvis, femur and tibia bones, knee joint, and foot structure--many of these changes being related to improved balance and shock absorption (eLucy 2007).
The morphological changes associated with bipedalism take millions of years to evolve. They first appear 6.0-7.0 million years ago (mya), but are not fully in place until around 4.0 mya. These physical changes continue to refine until we see them as we do today in modern Homo sapiens (Jurmain et al. 2013).
Hypotheses about the Evolution of Bipedalism
Several hypotheses have been proposed over the last century or so to explain the evolution of hominins. As bipedalism is the first trait on the road to modern humans, these hypotheses focus on the emergence of habitual bipedalism. Many have been refuted as new data is discovered. The first hypothesis was the hunting hypothesis proposed by Charles Darwin. The hunting hypothesis claims that the key to human evolution was the shift from an arboreal life to a terrestrial one. He predicted that the earliest hominins would be found in Africa based on the similarities he saw between humans and African apes. He suggested that bipedalism gave the first hominins an advantage in that it freed up their hands to carry weapons used to hunt animals. Darwin also suggested that larger brains preceded bipedalism as intelligence was needed to make the tools. Now we know that habitual bipedalism predates large brains so Darwin's hypothesis is no longer considered an adequate explanation. With the discovery of new data, other hypotheses have been proposed including the patchy-forest and provisioning hypotheses.
The patchy forest hypothesis suggests that the emerging mosaic environment that began at the end of the Miocene made bipedalism advantageous. The phrase mosaic environment in this case refers to an environment that had patchy forest interspersed with grasslands that eventually became the African savannas of today. This caused food resources to become spread out over the landscape. For traveling long distances, bipedalism is more energy efficient than quadrupedalism (walking on "all fours"). Traveling bipedally freed up hands for carrying provisions and the early hominins could have easily fed from both terrestrial and arboreal resources.
The provisioning hypothesis states that having hands free to carry food allowed males to provision females and offspring. Since much of the females energy went to child-rearing, the ability of a male to provision her and her offspring would have been an attractive quality. Those males who could walk more efficiently bipedally while carrying food would have been prime mate material, allowing both the male and female to reproduce successfully. However, species such as wolves provision their females and offspring even though they are quadrupeds by consuming meat and regurgitating it to the pups.
The truth of the matter is that the origins of bipedalism are still murky. Further research will hopefully help us come closer to a determination of why bipedalism, and hence our early ancestors, evolved. In the meantime, you can explore other hypotheses on the origins of bipedalism on the NOVA web site: http://www.pbs.org/wgbh/nova/evoluti...ipedalism.html [optional].
Dental Features
Apes have a chewing complex which is good for cutting and shredding food. Over time, hominins lose this dental feature, as the canine reduces in size, and the molars increase in size (Larsen 2014).
Brain Evolution
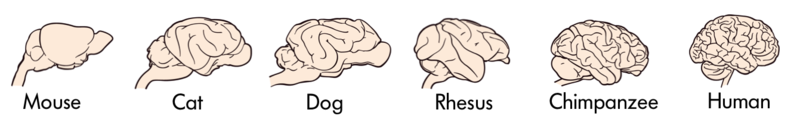
In relation to other mammals, primates have a more expanded and elaborate brain, including expansion of the cerebral cortex. Compare the complexity of the human brain on the left to the cat brain on the right (photos are not to scale).
Figure \(\PageIndex{6}\): Human Brain.Figure \(\PageIndex{7}\): Cat Brain.
Clearly, significant anatomical changes have taken place during the evolution of the brain in primates, in other mammals, and in animals in general. Are there generalizations that can be made about the evolution of animal brains?
Striedter (2006) has identified a number of general principles of brain evolution applicable across a wide range of species (i.e. not just primates or mammals).
1) Embryonic brains across species are more similar than adult brains, because brains tend to diversify more as they grow toward adult form;
2) relative brain size to body size in the vertebrates (animals with backbones) has tended to increase more often than decrease over evolutionary time;
3) it appears that increases in relative brain size were generally accompanied by increases in social or food foraging complexity;
4) most increases in relative brain size were accompanied by increases in absolute body size;
5) increases in absolute brain size require changes in the brain's internal connections which imply greater modularity or specialization (specialized processing modules increasing the "division of labor" as opposed to the whole brain doing all kinds of processing equally) of brain anatomy and functioning;
6) evolution generally enlarges brains by extending the period of brain development before and after birth while conserving (keeping the same) the "birth order" of different brain regions, so that big-brained animals tend to have disproportionately larger late-"born" (late-developing) regions ("late equals large"), such as cerebral cortex, leading to disproportionately more cerebral cortex (increased corticalization) in big-brained mammals (non-mammals don't have cerebral cortex); however there are exceptions to this rule, for example, at any given absolute brain size, there is more cerebral cortex in simians than in prosimians, and in parrots there is an unusually large telencephalon (forebrain) which is not accounted for by the rule;
7) changes in size proportions of brain areas, although "automatic" within the scaling (allometric) rules above, can still be adaptive and undergo natural selection;
8) as brain regions increase in absolute or proportional size, they tend to become laminated--organized into sheets of neurons--allowing point for point corresponding connections between sensory and motor maps with minimal axonal and dendritic wiring, saving space and metabolic energy;
9) as brain size increases, more regional subdivisions occur from ancestral parts subdividing into new parts, as in the dorsal thalamus (located below the cortex near the center of the brain), or, as in the case of neocortex, a new part was added onto an ancestral set of conserved (retained over evolution) brain parts;
10) a principle known as Deacon's rule is that "large equals well connected," meaning that as the relative size of a brain structure increases it tends to receive more connections and to project (send) more outputs to other structures.
Striedter (2006) adds a number of additional generalizations about the mammal and primate brains, including human brains:
11) six-layered mammalian neocortex (found only in mammals like us) probably evolved from a 3-layered reptilian precursor called dorsal cortex (something like that found in turtles) by addition of several layers of cortex;
12) aside from neocortex, the mammalian brain is similar to the reptilian brain (which also has hippocampus, for example) but even with a "fundamental scheme" of brain regions and circuitry, many minor changes in wiring can drastically change how information flows through a brain and thus how it functions--thus, the mammal brain is not just an upscale version of the reptilian brain;
13) increasing corticalization in mammals cannot be explained in terms of the above scaling (allometric) rules and involved highly specialized changes in brain anatomy presumably due to natural selection which expanded precursor sensory and motor cortical regions;
14) bird forebrains evolved along a very different path with expansion of their dorsal ventricular ridge (DVR), the major sensorimotor region of the avian telencephalon, highly similar in function to mammalian neocortex, making "many birds at least as intelligent as most mammals."
Striedter adds a number of points about the human brain in an attempt to identify features that make it special compared to the brains of other mammals.
15) In the six million years since bipedal apes (hominins) diverged from other apes, absolute brain size increased radically (about fourfold), not gradually, but in bursts--from when genus Homo first evolved, absolute brain size doubled from 400 to 800 cubic centimeters, then remained relatively steady in Homo erectus during the next 1.5 million years, but then exploded again in the transition to Homo sapien until about 100,000 years ago, at which time absolute brain size reached its current value of about 1,200 to 1,800 cubic centimeters. The first jump in Homo brain size was likely related to change in diet involving transition to meat and later the cooking of meat. The second leap was perhaps stimulated by competition among humans for mates and other resources;
16) the principle of "late equals large" predicts large neocortex in humans (the human neocortex to medulla ratio is twice that of chimpanzees);
17) the principle of "large equals well connected" is consistent with known expanded numbers of projections from human neocortex to motor neurons in medulla (located just above spinal cord) and spinal cord permitting greater precision of control over muscles serving hands, lips, tongue, face, jaw, respiratory muscles, and vocal folds, required for the development of human language about 50,000 to 100,000 years ago;
18) once human language appeared, dramatic changes in human behavior became possible without further increases in brain size;
19) increase in brain size has some disadvantages: including increased metabolic costs because the brain utilizes so much metabolic energy (20% of human metabolic energy even though it is only 2% of human body weight; being so metabolically expensive increases in brain size must be paid for by improved diet or reduction of other metabolic energy demands); decreased connectivity perhaps making the two hemispheres more independent of one another, and perhaps explaining why the two cerebral hemispheres became functionally specialized (performing different cognitive functions); and size limits of neonatal brain due to constraints imposed by size of the human mother's pelvis and birth canal. According to Striedter, these costs may explain why human brain size plateaued about 100,000 years ago;
20) within neocortex (found only in mammals), the lateral prefrontal cortex (located forward of your temples) has become relatively enlarged in the human brain, likely increasing its role in behavior (see Chapter 14 for additional discussion of the the lateral prefrontal cortex and higher cognitive functions such as thought, planning, etc.);
21) some key evolutionary changes in brain structure were not caused by increases in absolute or relative brain size, such as evolution of the neocortex of mammals, but require additional explanation; comparing distantly related species on absolute brain size alone misses important factors, for example, brains of some large whales weight 5 times as much as the human brain but whale brains have poorly laminated and thin neocortex; in cases of distantly related species, comparisons of relative brain size are more useful, for example, human and some toothed whales (e.g. killer whales) have relative brain sizes significantly larger than average mammals of similar body size;
22) two general hypotheses about brain evolution are that individual brain systems evolve independently by natural selection (the mosaic hypothesis) or alternatively that components of such systems evolve together because of functional constraints (the concerted or constraint hypothesis), with a third view being that all brain evolution is simultaneously both mosaic and concerted.
One problem with Striedter's approach is that it doesn't explicate the forces of natural selection that may account for more specific features of brain evolution in specific species, including our own. He admits that evolution of the neocortex cannot be explained by evolution of bigger brains and that neocortex evolved independently of absolute brain size. However, aside from restating the theory that complex social life in ancestral humans, along with competition among humans for resources, stimulated the evolution of increases in the size and complexity of the neocortex, he offers little insight about what role natural selection played in the evolution of the neocortex of mammals, or in brain evolution in general. As Adkins-Regan (2006, p.12-13) states in her critique of Striedter's work, "There is relatively little discussion of tests of hypotheses about the selective pressures responsible for the origin and maintenance of traits . . . The author would seem to be experiencing symptoms of discomfort with the concept of adaptation. . .. Given that brain mechanisms are products of natural selection, a central strategy in neuroscience should be to use the methods of evolutionary biology, which have been so successful in helping us to understand the mechanisms and design of organisms generally." In Chapter 14 of this text, on Intelligence and Cognition, consistent with the critique of Adkins-Regan, you will find an extensive discussion of the role of natural selection in evolution of brain systems involved in intelligence and thinking.
Encephalization of the Brain
Encephalization of the brain refers to a couple of things: 1) the increase in brain size over time and 2) the size of the brain in relation to total body mass. The brain-size to body mass ratio does not change that much in the hominins in spite of presumed increases in intelligence, however increases in brain size becomes significant beginning with the early Homo species. Studies of living mammal species suggest that increased number of neurons and increased density of neuron packing into the cranial cavity may be a key factor in the evolution of increased intelligence across species. However, while there is gradual increase in brain size throughout the australopithecine lineage, it is not until early Homo that there is a significant increase in cranial capacity, approximately a 20% increase over australopithecines. More significant is the approximately 50% increase in brain size of Homo erectus and the earlier Homo species. It is not just the size of the brain that is important. During this process of encephalization, it has been speculated, without fossil evidence, that there was also a rewiring of the brain that coincides with the emergence of material culture such as stone tools. It is not until this occurs that hominins leave Africa, enabled greatly by cultural advances.
Non-human primate brains are symmetrical as are the brains of early hominins. With the emergence of Homo we see the lateralization of the brain--it becomes asymmetrial (right brain, left brain). We know this from endocasts. Endocasts form when minerals replace brain matter inside the cranium during the fossilization process. These endocasts allow researchers to study the cortical folds of the brain and compare it to modern humans. Based on endocasts, researchers determined that three areas of the brain began to change in Homo: the cerebellum, which handles learned motor activities, the limbic system, which processes motivation, emotion and social communication, and the cerebral cortex, which is responsible for sensory experiences, memory, and complex mental functions such as language, cognition, planning, imagination, and intelligence (see chapters 14 and 15 on Cognition and Intelligence and Language). It is these changes that may have allowed the early members of our genus, Homo, to develop cultural adaptations to environmental pressures.
Figure \(\PageIndex{8}\): Comparison of cranial capacities of living primates. Primate skulls provided courtesy of the Museum of Comparative Zoology, Harvard University. (Image from Wikimedia Commons; File:Primate skull series with legend.png; https://commons.wikimedia.org/wiki/F...ith_legend.png; by Christopher Walsh, Harvard Medical School; licensed under the Creative CommonsAttribution-Share Alike 2.5 Generic license).
Why did the brain change in early Homo?
The question that confronted researchers was why the brain changed. Big brains have some disadvantages:
they require a lot of metabolic energy; approximately 25-30% of a human's metabolic energy is consumed by the brain although it comprises only 2% of total body weight
big brains require infants to be born in an immature state with head and brain size small enough to permit birth through a relatively small birth canal, resulting in a longer period of infant dependency (the average infant brain at birth is only about 1/3 the size of an adult brain with much of brain development and behavioral and mental development occurring over a period of years after birth)
longer infant dependency is an increased drain on maternal energy; the mother must have proper nutrition not only for herself but for the nursing infant
it has been suggested that larger brains decrease the bipedal efficiency of females because they must have a wider pelvis and birth canal to give birth to a large brained infant
So, for large brains to become evolutionarily fixed in the Homo population, the advantages had to outweigh the disadvantages listed above.
One possible explanation incorporates the interaction of three different variables: group size, complex subsistence patterns (such as foraging for food or domestication of animals and plants), and the nutritional value of meat (Campbell and Loy 2000: 318). Let's address group size first.
Research suggests that brain size and size of social groups correlate positively among living primates, implying that big brains helped individuals keep track of complex social information such as dominance hierarchies, alliances, enemies, etc.
Second, a big brain allows primates to keep track of large subsistence territories and allows omnivores to develop strategies for collecting a wide variety of foods.
Third, eating meat is a relatively easy way to get the nutrition needed to run a big brain, which, as mentioned above, in modern humans takes about 1/4 to 1/3 of our daily metabolic energy. However, raw meat requires a lot of energy to digest, so the invention of cooking meat over a fire may have played a role in evolution of larger brains because cooked meat is much easier, and requires less metabolic energy, to digest. Thus, cooking of meat provided a rich source of metabolic energy, supporting evolution of larger brains.
The argument for the social brain hypothesis is laid out by Robin Dunbar (1998). Dunbar also claims that it was changes in the neocortex, the 2-4 mm thick top layer of the cerebral hemispheres, that were critical in the "homininzation" (development of human cognitive abilities) of our ancestors (see chapter 14 for additional adaptive advantages associated with encephalization and increases in intelligence).
A related perspective on the evolution of the human brain emphasizes the role of culture in increasing brain size. As Geertz (2013, p. 180-181) states:
The Australopithecine brain was approximately the same size as that of modern-day chimpanzees (some 400– 600 cm3). The Homo sapiens brain is approximately 1200–1700 cm3. But the expansion of the brain had begun already with the appearance of Homo habilis, the first hominine species that came out of the Australopithecine line some 2.5 million years ago (with a brain size of 500–800 cm3) and became even more spectacular with the appearance 1.5 million years ago of the stone tool artist Homo erectus whose brain grew closer in size to modern humans at 750–1250 cm3. It is today assumed by archeologists and paleontologists that the production and use of tools was incremental to the expansion of the brain. Because tool use is much more than simply knocking stones and bones together, and indeed, it depends very much on the recipes, rules and instructions of cultural patterns, it is widely accepted today that culture drove the expansion of the brain (along with other things, of course, such as eating meat). The causal chain, once again, however, is culture first, brain expansion afterwards. . . . 'Because tool manufacture puts a premium on manual skill and foresight, its introduction must have acted to shift selection pressures so as to favor the rapid growth of the forebrain as, in all likelihood, did the advances in social organization, communication, and moral regulation which there is reason to believe also occurred during this period of overlap between cultural and biological change. Nor were such nervous system changes merely quantitative; alterations in the interconnections among neurons and their manner of functioning may have been of greater importance than the simple increase in their number.'
The Forebrain includes the cerebral cortex (frontal, parietal, temporal, and occipital lobes) and a number of subcortical structures such as the thalamus, hypothalamus, limbic system, and basal ganglia. Different selection pressures arising from tool manufacture and use, complex social organization, communication, moral regulation, and other social and cultural factors would have likely affected different areas of the forebrain, and even different lobes of the cerebral cortex, in different ways. This follows from the fact of brain organization that different brain circuits and networks are specialized for different cognitive and behavioral functions (see Cosmides & Tooby, 2002).
Geology and Environmental Background
The Miocene period (roughly 23-5 million years ago) was geologically active in Africa. This is the period of the adaptive radiation of the apes and a period of mountain building that led to the formation of the Great Africa Rift Valley. With the emergence of the rift mountains, the rains that heretofore had moved across the continent from the Atlantic Ocean were blocked (referred to as a rain shadow), leading to the aridification of Eastern Africa. The savanna environment that evolved in Eastern Africa was and is a much more open environment than the forested environment of Western and Central Africa, leading to rise of new adaptations for plants and animals. It is in this newly emerging environment that hominin evolution takes off.
Paleoclimatic data has been correlated with speciation events in hominin evolution, but it does not seem to account for all speciation events. Nonetheless, the paleoclimatic data suggests the following:
Grasslands spread in Africa between 10-5 million years ago during a cooling and drying phase. It is during this time frame that the common ancestor of African apes and humans lived. The common ancestor was more like a quadruped who was arboreal or at least spent a significant amount of time in the trees. In the middle of this period, approximately 7-6 million years ago, the first bipedal hominin emerged, and a few other early hominins are referred to as proto-hominins in recognition of their primitive, ape-like features.
In the mid-Pliocene period, 3-2 million years ago, yet another cooling and drying phase is correlated with the adaptive radiation of the hominins, including the emergence of the robust australopithecines in the genus Homo, the same genus as modern humans.
Near the beginning of the Pleistocene period, also known as the Ice Age, the environment continued to get drier. Open habitats spread in East Africa. During this period, Homo ergaster (Homo erectus) emerges and finally leaves the African continent.
This data has a tendency to make us think that hominin evolution was driven by environmental changes; however, geologic, climatic, and environmental changes in Africa during the Miocene, Pliocene and Pleistocene may have had little to do with the evolution of hominins, leaving open the possibility that the hypotheses discussed above, or others yet to be proposed, may be more causally related to the evolution of hominins.
Key Transitions in Human Evolution
On the evolutionary road to modern humans, some scientists have identified several "key transitions: (1) African ape to terrestrial bipedal ape (around 4 Ma)[Ma=million years ago]; (2) terrestrial bipedal ape (australopithecine) to ‘early Homo’ (around 2 Ma); (3) Early Homo [species] to Homo heidelbergensis (1–0.8Ma); (4) Homo heidelbergensis to larger-brained Homo (from 500 ka) [ka=thousand years ago], and (5) larger-brained Homo to H. sapiens (from 200 ka) . . . [Regarding social behavior,] "among archeologically based models, there is a total agreement that the emergence of basal hominin sociality, as part of our primate heritage, should be attributed to the very first evolutionary stages, possibly before the emergence of the Homo genus" (Anghelinu, 2013, p.13).
For Further Exploration
Explore Human Evolution in Print
Boyd, Robert and Joan B. Silk. 2009. How Humans Evolved, 5th edition. New York: W. W. Norton.
Campbell, Bernard G. and James D. Loy. 2000. Humankind Emerging, 8th edition. Boston: Allyn & Bacon.
Johanson, Donald and Kate Wong. 2010. Lucy's Legacy: The Quest for Human Origins. New York: Harmony Books.
Stringer, Chris and Peter Andrew. 2006. The Complete World of Human Evolution. New York: Thames & Hudson.
Tattersall, Ian. 2008. The Fossil Trail: How We Know What We Think We Know About Human Evolution. New York: Oxford University Press.
Explore Human Evolution on the Web
Becoming Human
Talk: Origins Fossil Hominids
Hall of Human Origins
Science Daily: Human Evolution News
Rediscovering Biology:Unit 9 Human Evolution
BBC: The Evolution of Man
Human Evolution: The Fossil Evidence in 3D
Proto-hominins
The oldest hominin discovered to date is from between 7.2 to 6.8 mya (million years ago).
Figure \(\PageIndex{9}\): Sahelanthropus tchadensis. The oldest known hominin.
The skull is a combination of ape-like and human-like features. Ape-like features include brain size, heavy brow ridge. Its human-like features include forward position of the foramen magnum, smaller canine teeth, and intermediate thickness of the premolar and molar enamel. Due to the pronounced brow ridge, Brunet’s team suggests that the specimen is male.
There is debate among researchers as to whether this fossil is a hominin or an ape. Some suggest that the specimen belongs to that of a female ape because it is likely to find canines worn at the tips in female apes.
Hominins
While the hominins will be presented more or less in chronological order, do not mistake chronological order for linear evolutionary relationships; some hominins that are presented are not in the direct line to modern humans. It is also important to keep in mind that new discoveries are made each year that refine what we know about human evolution.
Discovered in 1974 at Hadar, Ethiopia, Australopithecus afarensis is arguably the most well know fossil hominin species. It is dated from 3.7-3.0 mya (Scarre 2014). Over 40% of the skeleton was recovered, which allowed the team to fully reconstruct the skeleton. This fossil specimen, named Lucy, coupled with footprints found at Laetoli, Tanzania, in 1978 by Mary Leakey, confirmed that Au. afarensis was fully bipedal, albeit not exactly like modern humans. The footprints at Laetoli indicate that Au. afarensis had a short stride and a strolling gait. Since the 1970s, hundreds of specimens of Au. afarensis have been found (60 individuals at least from Hadar alone!) in Ethiopia, Kenya, and Tanzania, allowing paleoanthropologists to make “definitive statements about the locomotor pattern and stature of” (Jurmain 2013: 211) this early hominin.
Figure \(\PageIndex{10}\): Reconstruction of the fossil skeleton of "Lucy" the Australopithecus afarensis
Au. afarensis has several primitive, or ape-like, features, including a relatively small brain in comparison to Homo, a u-shaped dental arcade, a flat nose, a flattened forehead (referred to as platycephaly) and prognathic face (characterized by a protruding lower jaw). Its canines, while larger than Homo, are smaller than earlier hominins. While its brain was larger than earlier hominins, it is still small in comparison to genus Homo. There is evidence for sexual dimorphism; Au. afarensis males were no taller than 5 feet and females about 3-4 feet, similar in proportion to modern African apes. Au. afarensis has a suite of derived traits associated with bipedalism; i.e., bowl-shaped pelvis, s-curve of the vertebral column, and knee anatomy. However, the curvature of the fingers and toes and the proportion of the arms to legs suggest to some researchers that Au. afarensis spent some time in the trees.
In 2006, a 3.3 million year old Au. afarensis child was discovered less than 4 km from where Lucy was found in 1974. It is not only the oldest juvenile fossil ever found, it is also the most complete hominin fossil found to date. Selam, as the fossil was nicknamed (or Lucy's baby or Dikika's baby), confirmed earlier suggestions that Au. afarensis was bipedal, yet spent time in the trees. The shoulder structure with its upward pointing shoulder joints and the bony ridge running along the shoulder blades are like that of apes, which would have facilitated arboreal movement even if they were not as fully capable as chimpanzees at moving in trees.
Paleoecological data indicates that Au. afarensis lived in both grassland (savanna) and woodland environments.
Australopithecus africanus (see Figure 3.7.2, above) has been dated to 3-2 mya (Scarre 2014). In comparison to Au. afarensis, Au. africanus has smaller incisors and larger molars; the canines no longer have the pointed, triangular appearance seen in apes and Au. afarensis; however, Au. africanus still exhibits some protrusion of the lower jaw and has a small brain like that of Au. afarensis. There is general consensus that Au. africanus is a direct descendent of Au. afarensis. Its relationship to Homo is less clear.
Australopithecus garhi
Found in Bouri, Ethiopia in 1997, Au. garhi (see Figure 3.7.2, above) dates to 2.5 mya. Few fossil specimens have been found and those that have are relatively fragmentary. One cranium and other skull fragments were found and serve as the basis of the species identification. The size and shape of its molar teeth suggest to some researchers that Au. garhi is related to Paranthropus aethiopicus (see below), but its other features, e.g., braincase, face, and other teeth, are more like genus Homo. In light of this, some researchers contend it is ancestral to Homo.
Australopithecus sediba (Figure 3.3.13) was found in 2008 at Malapa Cave, South Africa, dating to 1.95-1.75 mya, has a mosaic of characteristics that suggest it may be transitional from the australopiths to genus Homo. However, this claim is controversial as the earliest dates for Homo predate Au. sediba by about 500,000 years (Becoming Human 2008).
The features that link Au. sediba to Homo include the pelvis shape, more vertical brain case, smaller cheek bones, and molar shape.
Robust Australopiths
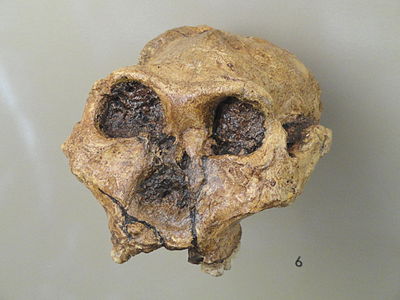
Three robust species of hominins emerged in the Plio-Pleistocene period: Paranthropus aethiopicus, Paranthropus boisei, and Paranthropus robustus (see Figures 3.3.14-17). They have morphological features that suggest they were well adapted for eating hard foods that needed grinding, which led to their being identified as “robust.”
Note
There is debate over whether the differences mentioned above qualify the robust australopiths to be in a separate genus from the australopithecines. In some anthropological works the genus Australopithecus is used. In others, such as this work, Paranthropus is used.
There is no evidence to suggest that P. boisei is ancestral to any subsequent hominin.
Figure \(\PageIndex{16}\). Paranthropus boisei; model of adult male (Smithsonian Museum of Natural History).
Figure \(\PageIndex{17}\). Paranthropus robustus.
Homo Genus
Above we described our oldest human ancestors, primarily members of the genus Australopithecus who lived between 2 million and 4 million years ago. Here we introduce the earliest members of the genus Homo, focusing on the species Homo habilis and Homo erectus (see Figure 3.3.19 and Figure 3.3.20, below).
Defining The Genus Homo
When grouping species into a common genus, biologists will consider criteria such as physical characteristics (morphology), evidence of recent common ancestry, and adaptive strategy (use of the environment). However, there is disagreement about which of those criteria should be prioritized, as well as how specific fossils should be interpreted in light of the criteria. There is general agreement that species classified as Homo should share characteristics broadly similar to our species. These include the following:
• a relatively large brain size, indicating a high degree of intelligence;
• a smaller and flatter face;
• smaller jaws and teeth; and
• increased reliance on culture, particularly the use of stone tools, to exploit a greater diversity of environments (adaptive zone).
Some researchers would include larger overall body size and limb proportions (longer legs/shorter arms) in this list. There is also an apparent decline in sexual dimorphism (body-size differences between males and females). While these criteria seem relatively clear-cut, evaluating them in the fossil record has proved more difficult, particularly for the earliest members of the genus. There are several reasons for this. First, many fossil specimens dating to this time period are incomplete and poorly preserved, making them difficult to evaluate. Second, early Homo fossils appear quite variable in brain size, facial features, and teeth and body size, and there is not yet consensus about how to best make sense of this diversity.
In this section, we will take several pathways toward examining the origin and evolution of the genus Homo. First, we will explore the environmental conditions of the Pleistocene epoch in which the genus Homo evolved. Next we will examine the fossil evidence for the two principal species traditionally identified as early Homo: Homo habilis and Homo erectus. Then we will use data from fossils and archeological sites to reconstruct the behavior of early members of Homo, including tool manufacture, subsistence practices, migratory patterns, and social structure. Finally, we will consider these together in an attempt to characterize the key adaptive strategies of early Homo and how they put our early ancestors on the trajectory that led to our own species, Homo sapiens.
Climate
The early hominin species covered previously, such as Ardipithecus ramidus and Australopithecus afarensis, evolved during the late Pliocene epoch. The Pliocene (5.3 million to 2.6 million years ago) was marked by cooler and drier conditions, with ice caps forming permanently at the poles. Still, Earth’s climate during the Pliocene was considerably warmer and wetter than at present.
The subsequent Pleistocene epoch (2.6 million years to 11,000 years ago) ushered in major environmental change. The Pleistocene is popularly referred to as the Ice Age. Since the term “Ice Age” tends to conjure up images of glaciers and woolly mammoths, one would naturally assume that this was a period of uniformly cold climate around the globe. But this is not actually the case. Instead, climate became much more variable, cycling abruptly between warm/wet (interglacial) and cold/dry (glacial) cycles.
In Africa, paleoclimate research has determined that grasslands expanded and shrank multiple times during this period, even as they expanded over the long term, becoming increasingly common during the Pleistocene. One solution adopted by some hominins was to specialize in feeding on the new types of plants growing in this landscape. The robust australopithecines probably developed their large molar teeth with thick enamel in order to exploit this particular dietary niche.
Members of the genus Homo took a different route. Faced with the unstable African climate and shifting landscape, they evolved bigger brains that enabled them to rely on cultural solutions such as crafting stone tools that opened up new foraging opportunities. This strategy of behavioral flexibility served them well during this unpredictable time and led to new innovations such as increased meat-eating, cooperative hunting, and the exploitation of new environments outside Africa, including Europe and Asia.
The Emergence of Homo (our Genus)
The emergence of the genus Homo marks the advent of larger brains, the emergence of material culture (at least material culture that survives in the archeological record), and the eventual colonization of the world outside of Africa. The earliest Homo species are contemporaneous with several australopiths: Au. africanus, Au. garhi, Au. sediba, and all of the Paranthropus species. Africa was flush with hominins (Figure \(\PageIndex{16}\)). There are several trends we see in the evolution of the earliest Homo species to humans, Homo sapiens:
Rounding of the cranium
Enlargement and rewiring of the brain (judged from endocasts)
Smaller faces and teeth
Decreasing prognathism
Tallness
Diversity of cultural traits
As with the overview on early hominins, we will continue taking the "lumper" approach - a tendency to lump specimens into fewer species rather than tending to differentiate and hypothesizing a greater number of species. Several Homo species will not be discussed in detail, but may be mentioned in passing, e.g., Homo rudolfensis, Homo ergaster, Homo gautengensis, Homo antecessor, Homo cepranensis, Homo rhodesiensis, Homo tsaichangensis. We will first examine the morphological characteristics of various Homo species. Cultural traits will be addressed separately.
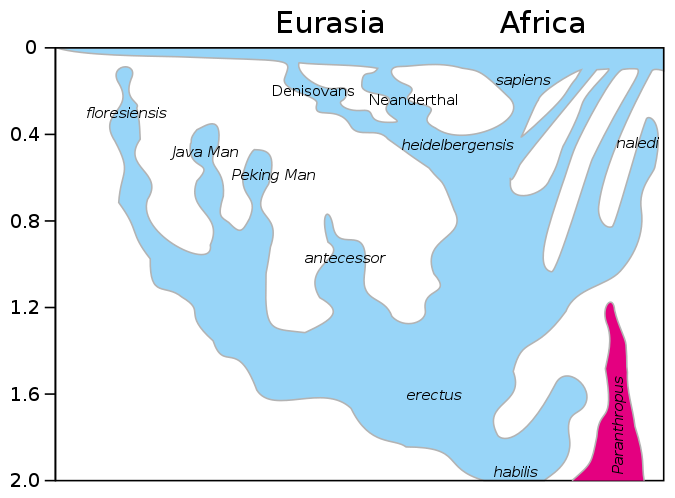
Figure \(\PageIndex{18}\): A model of the evolution of the genus Homo over the last 2 million years (millions of years, Mya, is on the vertical axis). The rapid "Out of Africa" expansion of H. sapiens is indicated at the top of the diagram by the lateral expansion of blue all the way across the top of the diagram, with admixture indicated with Neanderthals, Denisovans, and unspecified archaic African hominins. The late survival of robust australopithecines (Paranthropus) alongside Homo until 1.2 Mya is indicated in purple.
H. heidelbergensis is shown as the link between Neanderthals, Denosiovans and H. sapiens
Division of Asian H. erectus into Java Man and Peking Man
H. antecessor shown as a branch of H. erectus reaching Europe
After H. sapiens emerge from Africa some 60 kya they spread across the globe and interbred with other descendants of H. heidelbergensis, and Neanderthals, Denisovans
Homo habilis has traditionally been considered the earliest species placed in the genus Homo. However, as we will see, there is substantial disagreement among paleoanthropologists about the fossils classified as Homo habilis, including whether they come from a single or multiple species, or even whether they should be part of the genus Homo at all.
Compared to the australopithecines, Homo habilis has a somewhat larger brain size–an average of 650 cubic centimeters (cc) compared to less than 500 cc for Australopithecus. Additionally, the skull is more rounded and the face less prognathic. However, the postcranial remains show a body size and proportions similar to Australopithecus. Known dates for fossils identified as Homo habilis range from about 2.5 million years ago to 1.7 million years ago.
Dates 2.5 million years ago to 1.7 million years ago
Region(s): East and South Africa
Famous Discoveries: Olduvai Gorge, Tanzania; Koobi Fora, Kenya; Sterkfontein, South Africa
Brain Size: 650 cc average (range from 510 cc to 775 cc)
Dentition: Smaller teeth with thinner enamel compared to Australopithecus; parabolic dental arcade shape
Cranial Features: Rounder cranium and less facial prognathism than Australopithecus
Postcranial Features: Small stature; similar body plan to Australopithecus
Culture: Oldowan tools; oldest of known stone tool type; scrapers, choppers; indication of changing cognitive abilities (see Module 3.8)
Stone tools almost certainly predated Homo habilis (possibly by Australopithecus garhi or the species responsible for the tools from Kenya dating to 3.7 million years ago). However, stone tools become more frequent at sites dating to about 2 million years ago, the time of Homo habilis (Roche, Blumenschine, and Shea 2009). This suggests that these hominins were increasingly reliant on stone tools to make a living.
Stone tools are assigned a good deal of importance in the study of human origins. Studying the form of the tools, the raw materials selected, and how they were made and used can provide insight into the thought processes of early humans and how they modified their environment in order to survive (see Supplementary Content, Chapter 18, Material Culture). Paleoanthropologists have traditionally classified collections of stone tools into industries, based on their form and mode of manufacture. There is not an exact correspondence between a tool industry and a hominin species; however, some general associations can be made between tool industries and particular hominins, locations, and time periods. The names for the four primary tool industries in human evolution (from oldest to most recent) are the Oldowan, Acheulean, Mousterian, and Upper Paleolithic. The oldest stone tool industry is the Oldowan, named after the site of Olduvai Gorge where the tools were first discovered. The time period of the Oldowan is generally considered to last from about 2.5 mya to 1.6 mya. The tools of this industry are described as “flake and chopper” tools—the choppers consisting of stone cobbles with a few flakes struck off them. To a casual observer, these tools might not look much different from randomly broken rocks. However, they are harder to make than their crude appearance suggests. The rock selected as the core must be struck by the rock serving as a hammerstone at just the right angle so that one or more flat flakes are removed. This requires selecting rocks that will fracture predictably instead of chunking, as well as the ability to plan ahead and envision the steps needed to create the finished product. The process leaves both the core and the flakes with sharp cutting edges that can be used for a variety of purposes.
What were the hominins doing with the tools? One key activity seems to have been butchering animals. Animal bones with cutmarks start appearing at sites with Oldowan tools. Studies of animal bones at the site show leg bones are often cracked open, suggesting that they were extracting the marrow from the bone cavities. It is interesting to consider whether the hominins hunted these animals or acquired them through other means. The butchered bones come from a variety of African mammals, ranging from small antelope to animals as big as wildebeest and elephants! It is difficult to envision slow, small-bodied Homo habilis with their Oldowan tools bringing down such large animals. One possibility is that the hominins were scavenging carcasses from lions and other large cats. Regardless of how they were acquiring the meat, all these activities suggest an important dietary shift from the way that the australopithecines were eating. The Oldowan toolmakers were exploiting a new ecological niche that provided them with more protein and calories. Overall, increasing use of stone tools allowed hominins to expand their ecological niche and exert more control over their environment. As we’ll see shortly, this pattern continued and became more pronounced with Homo erectus.
Discovery of Homo habilis
Homo habilis was first discovered by Louis and Mary Leakey at Olduvai Gorge, Tanzania in 1960. Associated with stone tools (Oldowan), the Leakeys named their discovery “handy man.” H. habilis fossils have been found in Tanzania, Kenya, Ethiopia, and South Africa, although there is some debate as to whether the South Africa specimens should be included in the species. Some researchers contend that there was another early Homo species, Homo rudolfensis, which dates back to 2.4-2.5 mya. The H. rudolfensis fossils are slightly larger than those of H. habilis, leading some researchers to suggest the H. habilis exhibited sexual dimorphism and what we are seeing are male and female specimens of H. habilis. Others claim the size differences are significant enough to warrant the two species designations (O’Neil 1999-2012). In 2013 a Homo mandible was discovered in the Ledi-Geraru research area, Afar, Ethiopia. Dated to 2.8 to 2.75mya, the mandible exhibits an Australopithecus-like chin and Homo-like teeth (Villmoare et al. 2015). While still early in the research process, this discovery and further research may push back the date of the origin of Homo and help to resolve the debate between the H. rudolfensis and H. habilis fossils. For our purpose, we will consider them all H. habilis, making the approximate date range for this hominin 2.5 to 1.4mya or less conservatively to 1.7mya.
Figure \(\PageIndex{19}\): Homo habilis.
Morphologically, H. habilis has a larger brain than the australopiths, about 35% larger (O’Neil c1999-2012). You will recall from the section on trends in human evolution that it is speculated that the brain also began to rewire at this point. H. erectus exhibits less prognathism (protrusion of the lower jaw) and platycephaly (flattening of the back of the head) than early hominins. The brow ridge is also smaller. All of these traits together make the face smaller than the australopiths. Postcranially, H. habilis exhibit a mix of primitive and derived traits. Primitive traits connecting it to an australopith ancestor are the longer forearms and the size of the finger bones along with how the tendons attach to the wrist bones. The tips of the finger bones are broad like humans. Smaller teeth, a dental arcade in the shape of a parabolic arch, foot morphology, and a more rounded skull complete the human-like traits. Microanalysis of tooth wear indicates that H. habilis was omnivorous.
Table 3.3.2
Key Homo habilis fossil locations and the corresponding fossils and dates(adapted by Kenneth A. Koenigshofer, PhD, Chaffey College, from Explorations: An Open Invitation to Biological Anthropology; https://explorations.americananthro.....php/chapters/; Chapter 10, "Early Members of the Genus Homo" by Bonnie Yoshida-Levine Ph.D.; Beth Shook, Katie Nelson, Kelsie Aguilera, and Lara Braff, Eds.; licensed under a Creative Commons Attribution-NonCommercial 4.0 International License, except where otherwise noted).
Location of Fossils Dates (mya = millions of years ago) Description
Ledi-Gararu, Ethiopia 2.8 mya Partial lower jaw with evidence of both Australopithecus and Homo traits; tentatively considered oldest Early Homo fossil evidence.
Olduvai Gorge, Tanzania 1.7 mya to 1.8 mya. Several different specimens classified as Homo habilis, including the type specimen found by Leakey, a relatively complete foot, and a skull with a cranial capacity of about 600 cc.
Koobi Fora, Lake Turkana Basin, Kenya 1.9 mya. Several fossils from Lake Turkana basin show considerable size differences, leading some experts to classify the larger specimen as a separate species, Homo rudolfensis.
Sterkfontein and other possible about 1.7 mya South African caves have yielded fragmentary remains identified as Homo habilis, but South African cave sites secure dates and specifics about the fossils are lacking.
Homo erectus
About 1.9 mya, a new species of Homo appeared. Known as Homo erectus, the prevailing scientific view was that this species was much more like us. These hominins were equipped with bigger brains and large bodies with limb proportions similar to our own. Perhaps most importantly, their way of life is now one that is recognizably human, with more advanced tools, hunting, use of fire, and colonizing new environments outside of Africa.
Compared to Homo habilis, Homo erectus had a larger brain size (average of about 900 cc compared to 650 cc to 750 cc for habilis). Instead of having a rounded shape like our skulls have, the erectus skull was long and low like a football, with a receding forehead, and a horizontal ridge called an occipital torus that gave the back of the skull a squared-off appearance. The cranial bones are thicker than those of modern humans, and some Homo erectus skulls have a slight thickening along the sagittal suture called a sagittal keel. Large, shelf-like brow ridges hang over the eyes. As noted above, the climate was increasingly arid and the forest canopy in parts of Africa was being replaced with a more open grassland environment, resulting in increased sun exposure for our ancestors. Compared to the earlier australopithecines, members of the genus Homo were also developing larger bodies and brains, starting to obtain meat by hunting or scavenging carcasses, and crafting sophisticated stone tools. For purposes of cooling, H. erectus may have had little body hair accompanied by darkened skin to protect against sun exposure. It is generally agreed that Homo erectus was the first hominin to migrate out of Africa and colonize Asia and later Europe (although recent discoveries in Asia may challenge this view).
Homo erectus Fossils
Based on current fossil data, Homo erectus existed between 1.9 million (mya) to 25 thousand years ago (Jurmain 2013). H. erectus, literally “upright human,” fossils have been found in Java (Indonesia), Africa, China, Europe, and Israel. Based on morphological differences in the cranium, some scientists identify two species, H. erectus in Asia and H. ergaster in Africa with the African specimens being smaller than the Asian; however, we will use the H. erectus designation for both.
Figure \(\PageIndex{20}\): Homo erectus, Turkana Boy.
In 1984, a nearly complete H. erectus skeleton was found along a river in northern Kenya. Potassium-argon dating places “Turkana Boy” between 1.64 and 1.33 million years ago (mya). Aging and sexing of the fossil remains indicate that the individual was a male about age eight. He stood about 5’3” tall. Recent studies indicate that Turkana Boy followed the growth pattern of apes, so would have been near his adult height at the time of his death (Jurmain et al. 2013).
Homo erectus has a long history in Indonesia; fossils from Java were dated by argon dating to about 1.6 million to 1.8 million years. H. erectus fossils from the site of Ngandong in Java have yielded very recent dates of 43,000 years, although a more recent study with different dating methods concluded that they were much older—between 140,000 and 500,000 years old.
The pattern of increased brain size continued with H. erectus; its brain is up to 50% larger than its predecessor, H. habilis (O’Neil c1999-2012). This large brain was supported by a diet heavy in meat and other proteins. Its distinguishing characteristics include its sagittal keeling (a thickening of bone that runs from front to back on top of the skull; likely for muscle attachment) massive brow ridges (supraorbital tori) and bony prominences on the back of the skull. Postcranially, H. erectus’ bones are thicker than H. habilis, as were its jaws and face bones, and the proportion of arms to legs is like that of modern humans, causing some to suggest that its bipedal gait was like ours. The length of its leg bones indicates that H. erectus would have been an efficient long-distance runner, allowing them to “run down small and even medium size game animals on the tropical savannas of East Africa” (O’Neil c1999-2012). If so, as mentioned above, it is likely that H. erectus had much less body hair than its predecessors, as they would have needed to be able to sweat efficiently to cool the body. It is possible that H. erectus had little body hair (NOVA 2011). During the Pleistocene, the climate was increasingly arid and the forest canopy in parts of Africa was being replaced with a more open grassland environment, resulting in increased sun exposure for our ancestors.
Compared to the earlier australopithecines, members of the genus Homo were also developing larger bodies and brains, starting to obtain meat by hunting or scavenging carcasses, and crafting sophisticated stone tools. According to Nina Jablonski, an expert on the evolution of human skin, the loss of body hair and increased sweating capacity are part of the package of traits characterizing the genus Homo. While larger brains and long-legged bodies made it possible for humans to cover long distances while foraging, this new body form had to cool itself effectively to handle a more active lifestyle. Preventing the brain from overheating was especially critical. The ability to keep cool may have also enabled hominins to forage during the hottest part of the day, giving them an advantage over savanna predators, like lions, that typically rest during the heat of the day.
As noted above, scientists generally agree that H. erectus was the first hominin to leave Africa. As mentioned previously, fossils have been found in Africa, northern China, Indonesia, Europe, and Israel. In the Republic of Georgia, fossils were found in strata dated to 1.7mya suggesting that H. erectus left Africa soon after it evolved. A recent report (Dembo et al. 2015) posits that H. habilis was the first hominin to leave Africa, not H. erectus. Should this contention be supported with more data, it can still be argued that H. erectus was quite successful in colonizing the Old World (Africa, Europe, and Asia), helped, no doubt, by its advanced cultural behaviors.
Now we can address the question of why Homo erectus traveled such vast distances to these far-flung regions. To do this, we have to consider what we have learned about the biology, culture, and environmental circumstances of Homo erectus. The larger brain and body size of Homo erectus were fueled by a diet consisting of more meat, and longer more powerful legs made it possible to walk and run longer distances to acquire food. Since they were eating higher on the food chain, it was necessary for them to extend their home range to find sufficient game. Cultural developments including better stone tools and new technology such as fire gave them greater flexibility in adapting to different environments. Finally, the major Pleistocene climate shift discussed earlier in the chapter certainly played a role. Changes in air temperature, precipitation, access to water sources, and other habitat alteration had far-reaching effects on animal and plant communities; this included Homo erectus. If hominins were relying more on hunting, the migration patterns of their prey could have led them increasingly long distances.
There is evidence of Homo erectus in China from several regions and time periods. Homo erectus fossils from northern China, collectively known as “Peking Man,” are some of the most famous human fossils in the world, dated to about 400,000–700,000 years ago. The discovery of stone tools from China dating to 2.1 million years, older than any Homo erectus fossils anywhere, opens up the intriguing possibility that hominins earlier than Homo erectus could have migrated out of Africa.
At this time, researchers aren’t in agreement as to whether the first Europeans belonged to Homo erectus proper or to a later descendent species.
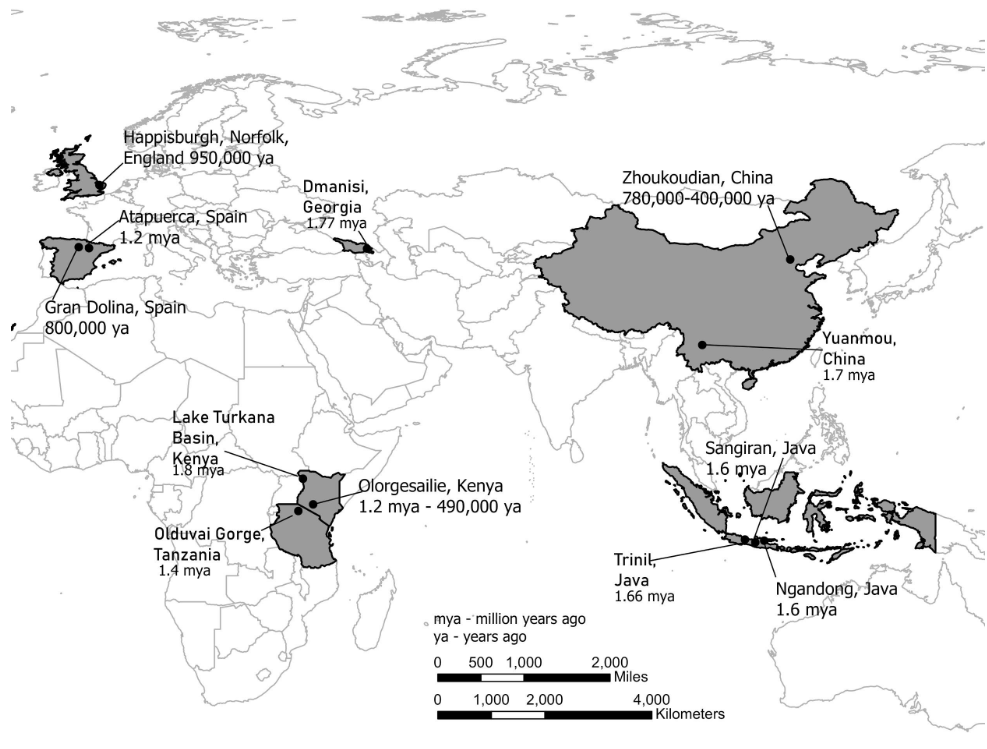
Figure \(\PageIndex{21}\): Map showing the locations of Homo erectus fossils around Africa and Eurasia (adapted by Kenneth A. Koenigshofer, PhD, Chaffey College, from Explorations: An Open Invitation to Biological Anthropology; https://explorations.americananthro.....php/chapters/; Chapter 10, "Early Members of the Genus Homo" by Bonnie Yoshida-Levine Ph.D.; Beth Shook, Katie Nelson, Kelsie Aguilera, and Lara Braff, Eds.; licensed under a Creative Commons Attribution-NonCommercial 4.0 International License, except where otherwise noted).
Homo erectus Culture
Homo erectus shows significant cultural innovations in diet, technology, life history, environments occupied, and perhaps even social organization, some that you will probably recognize as more “human-like” than any of the hominins previously covered. About 1.5 million years ago, some Homo erectus populations began making different forms of tools (known as "material culture;" see Section 3.14). These tools–classified together as constituting the Acheulean tool industry–are more complex in form and more consistent in their manufacture. Unlike the Oldowan tools, which were cobbles modified by striking off a few flakes, Acheulean toolmakers carefully shaped both sides of the tool. This type of technique, known as bifacial flaking, requires more planning and skill on the part of the toolmaker; he or she would need to be aware of principles of symmetry when crafting the tool. One of the most common tool forms of this period was the handaxe. Besides handaxes, forms such as scrapers, cleavers, and flake tools are present at Homo erectus sites. One striking aspect of Acheulean tools is their uniformity. They are more standardized in form and mode of manufacture than the earlier Oldowan tools. For example, the aforementioned handaxes vary in size, but they are remarkably consistent in regard to their shape and proportions. They were also an incredibly stable tool form over time—lasting well over a million years with little change.
Recently, newer methods—including microscopic analysis of burned rock and bone—have revealed clear evidence of fire use at Koobi Fora, Kenya, dating to 1.5 million years ago (Hlubik et al. 2017).
There is general consensus that H. erectus evolved from H. habilis and Homo heidelbergensis evolved from H. erectus in Africa, eventually supplanting H. erectus populations in the Old World--Africa, Asia, and Europe (Figure \(\PageIndex{18}\)).
Tool Use and Cognitive Abilities of Homo erectus
What (if anything) do the Acheulean tools tell us about the mind of Homo erectus? Clearly, they took a fair amount of skill to manufacture. Apart from the actual shaping of the tool, other decisions made by toolmakers can reveal their use of foresight and planning. Did they just pick the most convenient rocks to make their tools, or did they search out a particular raw material that would be ideal for a particular tool? Analysis of Acheulean stone tools suggest that at some sites, the toolmakers selected their raw materials carefully—traveling to particular rock outcrops to quarry stones and perhaps even removing large slabs of rock at the quarries to get at the most desirable material. Such complex activities would require advanced planning. They also likely required cooperation and communication with other individuals, as such actions would be difficult to carry out solo. However, other Homo erectus sites lack evidence of such selectivity; instead of traveling even a short distance for better raw material, the hominins tended to use what was available in their immediate area (Shipton et al. 2018). In contrast to Homo erectus tools, the tools of early modern Homo sapiens during the Upper Paleolithic display tremendous diversity across regions and time periods. Additionally, Upper Paleolithic tools and artifacts communicate information such as status and group membership. Such innovation and social signaling seem to have been absent in Homo erectus, suggesting that they had a different relationship with their tools than did Homo sapiens (Coolidge and Wynn 2017). Some scientists assert that these contrasts in tool form and manufacture may signify key cognitive differences between the species, such as the ability to use a complex language.
Table 3.3.3. Characteristics ofHomo erectus. (adapted by Kenneth A. Koenigshofer, PhD, Chaffey College, from Explorations: An Open Invitation to Biological Anthropology; https://explorations.americananthro.....php/chapters/; Chapter 10, "Early Members of the Genus Homo" by Bonnie Yoshida-Levine Ph.D.; Beth Shook, Katie Nelson, Kelsie Aguilera, and Lara Braff, Eds.; licensed under a Creative Commons Attribution-NonCommercial 4.0 International License, except where otherwise noted).
Hominin
Homo erectus
Dates
1.8 million years ago to about 200,000 years ago
Region(s)
East and South Africa; West Eurasia; China and Southeast Asia
Famous Discoveries
Lake Turkana, Olorgesailie, Kenya; Zhoukoudian, China; Dmanisi, Republic of Georgia
Brain Size
Average 900 cc; range between 650 cc and 1,100 cc
Dentition
Smaller teeth than Homo habilis
Cranial Features
Long, low skull with robust features including thick cranial vault bones and large brow ridge, sagittal keel, and occipital torus
Postcranial Features
Larger body size compared to Homo habilis; body proportions (longer legs and shorter arms) similar to Homo sapiens
Culture
Acheulean tools (in Africa) (see Module 3.8); evidence of increased hunting and meat-eating; use of fire; migration out of Africa
THE BIG PICTURE OF EARLY HOMO
We are discovering that the evolution of the genus Homo is more complex than what was previously thought. The earlier prevailing view of a simple progression from Australopithecus to Homo habilis to Homo erectus as clearly delineated stages in human evolution just doesn’t hold up anymore.
Variability in the Fossil Record of Early Homo
As is apparent from the information presented here, there is tremendous variability during this time. While fossils classified as Homo habilis show many of the characteristics of the genus Homo, such as brain expansion and smaller tooth size, the small body size and long arms are more akin to australopithecines. There is also tremendous variability within the fossils assigned to Homo habilis, so there is no consensus on whether it is a single or multiple species of Homo, a member of the genus Australopithecus, or even a yet-to-be-defined new genus.
What does this diversity mean for how we should view early Homo? First, there isn’t an abrupt break between Australopithecus and Homo habilis or even between Homo habilis and Homo erectus. Characteristics we define as Homo don’t appear as a unified package; they appear in the fossil record at different times. This is known as mosaic evolution.
We can consider several explanations for the diversity we see within early Homo from about 2.5 million to 1.5 million years ago. One possibility is the existence of multiple contemporaneous species of early Homo during this period. In light of the pattern of environmental instability discussed earlier, it shouldn’t be surprising to see fossils from different parts of Africa and Eurasia display tremendous variability. Multiple hominin forms could also evolve in the same region, as they diversified in order to occupy different ecological niches. However, even the presence of multiple species of hominin does not preclude their interacting and interbreeding with one another.
Diversity of brain and body sizes could also reflect developmental plasticity—short-term adaptations within a lifetime (Anton, Potts, and Aiello, 2014). These have the advantage of being more flexible than genetic natural selection, which could only occur over many generations. For example, among human populations today, different body sizes are thought to be adaptations to different climate or nutritional environments.
Trends in the Behavior of Early Homo
New discoveries are also questioning old assumptions about the behavior of Homo habilis and Homo erectus. Just as the fossil evidence doesn’t neatly separate Australopithecus and Homo, evidence of the lifeways of early Homo show similar diversity. For example, one of the traditional dividing lines between Homo and Australopithecus was thought to be stone tools: Homo made them; Australopithecus didn’t. However, the recent discovery of stone tools from Kenya dating to 3.3 million years ago challenges this point of view. Similarly, the belief that Homo erectus was the first species to settle outside Africa may now come into question with the report of 2.1 million-year-old stone tools from China. If this find is supported by additional evidence, it may cause a reevaluation of Homo erectus being first to leave. Instead, there could have been multiple earlier migrations of hominins such as Homo habilis or even Australopithecus species.
Rather than obvious demarcations between species and their corresponding behavioral advancements, it now looks like many behaviors were shared among species. Despite the haziness dominating the early Homo narrative, we can identify some overall trends for the million-year period associated with early Homo. These trends include brain expansion, a reduction in facial prognathism, smaller jaw and tooth size, larger body size, and evidence of full terrestrial bipedalism. These traits are associated with a key behavioral shift that emphasizes culture as a flexible strategy to adapt to unpredictable environmental circumstances. Included in this repertoire are the creation and use of stone tools to process meat obtained by scavenging and later hunting, a utilization of fire and cooking, and the roots of the human life history pattern of prolonged childhood, cooperation in child raising, and the practice of skilled foraging techniques. In fact, it’s apparent that the cultural innovations are driving the biological changes, and vice versa, fueling a feedback loop that will continue during the later stages of human evolution.
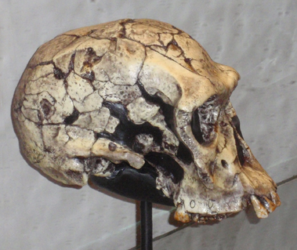
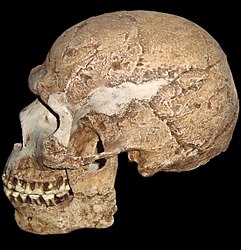
Homo heidelbergensis : the common ancestor of Homo neanderthalensis in Europe and Homo sapiens in Africa.
Some publications, e.g. Larsen 2014, refer to H. heidelbergensis as archaic Homo sapiens, but for our purposes, we will use the H. heidelbergensis designation. Otto Schoetensack found the first Homo heidelbergensis fossils in 1907 in Mauer, Germany. Since then H. heidelbergensis fossils have been found in Africa, Europe, and Asia. The date range for the species is 800 kya (thousand years ago) to 350 kya. Primitive traits (traits like the ancestors) include its large supraorbital tori (brow ridge), sagittal keeling, and low frontal bone. Derived traits (traits different from the ancestor) include separate supraorbital tori over each eye orbit, a more vertical posterior cranial vault, wide parietal bones in relation to the cranial base, and a larger cranial capacity than H. erectus (Becoming Human c2008). Additionally, they exhibit sexual dimorphism similar to that of modern humans.
Figure \(\PageIndex{22}\): Homo heidelbergensis skull on mirrored steel stand showing reflection of palate and dentition in upper jaw.
There is regional variation in the morphology of H. heidelbergensis. European specimens found at Atapuerca, (Spain), Petralona (Greece), Seinheim (Germany), and Swancombe (England) show that they had compact bodies, which could have been a response to living in the cold climates of the north as it would help to conserve heat. Additionally, the cranium is a mosaic of H. erectus traits and derived traits. In Asia, data from sites such as Zhoukoudian, Jinniushan, and Dali (China) show a mix of H. erectus and H. sapiens traits; the latter includes large cranial capacity and thin braincase walls. African specimens from Kabwe (Zambia), Florisbad (South Africa), Laetoli (Tanzania), and Bodo (Ethiopia) also show a combination of H. erectus and H. sapiens traits. It shares the massive supraorbital tori and prominent occipital torus with H. erectus and thin cranial vault bones, less angulated occipital, and cranial base with H. sapiens.
Significantly, H. heidelbergensis is the common ancestor of Homo neanderthalensis in Europe and Homo sapiens in Africa.
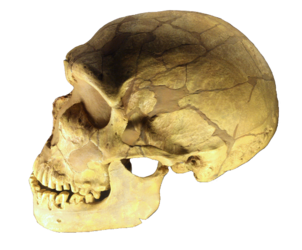
Homo neanderthalensis
Numerous Neanderthal fossils have been recovered since its discovery in 1856 in the Neander Valley, Germany. Neanderthals have been the speculation of scientists and the general public ever since. Some anthropologists classify Neanderthals as a subspecies of Homo sapiens, Homo sapiens neanderthalensis, while others interpret the morphological differences as significant enough to warrant classifying them as a different species, Homo neanderthalensis. Here, we will use the latter designation.
Figure \(\PageIndex{23}\): Homo neanderthalensis.
True Neanderthals first appear in the fossil record about 200,000 years ago (200 kya), with fossils exhibiting Neanderthal-like characteristics appearing as early as 400 kya. Recent research indicates that Neanderthals went extinct only about 40,000 years ago, between 41 kya and 39 kya (Higham et al., 2014). Molecular research denotes that some Neanderthal DNA lives on in modern humans, approximately 2% of the DNA of “people who descend from Europeans, Asians, and other non-Africans is Neanderthal” (Callaway, 2014). The Neanderthal genes are involved in fighting infections, dealing with ultraviolet radiation (Callaway, 2014), and living at high altitudes (Callaway, 2015). Neanderthal DNA has recently been linked with depression, obesity, and certain skin disorders, e.g., lesions caused by sun exposure (Callaway, 2015).
Neanderthals are the only hominin to originally evolve in a glacial environment, leading to some characteristics adapted for the cold climate. Many scientists contend that the midface prognathism (protruding midface) allowed for enlarged sinuses that functioned to warm and add moisture to the cold, dry air before entering the lungs. Small holes below the eye orbits, called the infraorbital foramina, are larger in the European Neanderthals that modern humans, suggesting that the blood vessels were larger, which allowed for more blood flow to the face. This would have helped keep the face warmer. They are relatively short and stocky, males averaging 5’ 5” and females 5’1”, with shorter appendages, both of which would help to conserve heat by providing less surface area from which to radiate heat. Their leg bones are thick and dense, suggesting that they frequently walked and ran, most likely in food procurement activities. Some postulate that some Neanderthals had pale skin that would have helped increase Vitamin D synthesis by increasing the amount of UV radiation to be absorbed by the body (O’Neil, c1999-2014). The width of their body trunks and short tibias fit the predictions for cold weather adaptation as proposed by Christopher Ruff (Larsen, 2013). The brain size of Neanderthals may also be related to cold weather. Averaging a cranial capacity between 1300 to 1400 cm3, they have the largest brains of all hominins, including our own species, Homo sapiens. The size may be associated with increased metabolic efficiency in cold weather, which is similar to modern Inuit peoples today who have a larger brain size than other human populations (Jurmain et al., 2013).
The presence of an occipital bun (a prominent bulge at the lower back of the skull) is one of the characteristics used to identify H. neanderthalensis specimens, although it should be noted that this characteristics persists in a small percentage of modern human populations. The occipital bun may have evolved to counterbalance Neanderthal’s heavy face when running; it prevents the head from making huge horizontal accelerations (NOVA 2002). The occipital bun also makes the skull elongated in comparison to most Homo sapiens. They had heavy brow ridges like those of H. heidelbergensis. Neanderthal skulls recovered from Amud and Tabun in Israel exhibit more H. sapiens-like cranial characteristics, including lack of an occipital bun, smaller eye orbits, tall and wide nasal openings, and smaller teeth (Larsen 2014).
Denisovans
The Denisovans were archaic humans closely related to Neanderthals, whose populations overlapped geographically with the ancestors of modern-day humans who existed at the same time. In 2010, scientists announced the discovery and DNA analysis of a finger bone and two teeth found in Denisova Cave, Siberia (Reich et al., 2010). The artifacts were recovered from a deposit dated to 50 kya to 30 kya. Data suggest that the remains were from an individual who shared a common origin with Neanderthals, but was not a Neanderthal, nor was it a modern human. The Denisovan individual(s) share 4-6% of its genetic material with modern peoples living in New Guinea, Bougainville Islands, and China. Further studies (Cooper and Stringer, 2013) indicate that the Denisovans crossed Eurasia and interbred with modern humans, but genetically the Denisovans were more closely related to Neanderthals than modern humans (Meyer et al., 2012). While further finds will shed more light on the Denisovans, it is clear that there was more genetic variability during the Pleistocene than previously thought (Larsen, 2014), but that gene flow between the various populations had implications for the emergence of modern humans (Pääbo, 2015).
Figure \(\PageIndex{24}\): Spread and Evolution of Denisovans.
Homo floresiensis
In 2003, a team of scientists discovered unusual fossils in Liang Bua cave, Flores Island, Indonesia. The small stature of the individual led to the naming of a new species, Homo floresiensis. Since then twelve individuals have been found ranging in time from 74 kya to 17 kya. H. floresiensis had a small brain and stood only about 3.5’ tall as an adult. They had receding foreheads, no chins, shrugged-forward shoulders, and large feet relative to their short legs. They share some characteristics with H. sapiens, including smaller dentition, separate brow ridges, and a non-prognathic face (flattened face).
Figure \(\PageIndex{25}\): Homo floresiensis.
Several hypotheses have been proposed to explain the appearance of H. floresiensis. One suggests that we are seeing island dwarfism, which is an evolutionary process resulting from long-term isolation on a small island. Limited resources and lack of predators selects for smaller bodied individuals who need fewer resources then large-bodied individuals. Another hypothesis claims that H. floresiensisis is not a separate species, but H. sapiens exhibiting microcephaly or some other developmental deficiency such as hyperthyroid cretinism (Oxnard et al. 2010) because the “…cranial features…are within the modern range of variation seen in living populations from the larger region [of Indonesia]” (Larsen 2014: 409), lending support to the second hypothesis. How H. floresiensis fits into the evolutionary picture is unclear based on the current data.
Homo sapiens
At present, only one hominin species inhabits planet Earth, Homo sapiens. All other Homo species became extinct by approximately 30,000 to 40,000 years ago. Many scientists believe that these extinctions were driven by competition from Homo sapiens. Morphologically, H. sapiens characteristics include the presence of a chin, a large brain, flat face, rounded or globular cranium, and a continuous, reduced brow ridge. Its bones are gracile (slender) in comparison to earlier hominins, although the earliest H. sapiens are more robust than modern populations and show none of the cold weather adaptations of Neanderthals. The more gracile nature lends credence to the hypothesis that modern humans evolved first in Africa. The leaner body proportions are more adaptive to the tropical African environments as there would be more body surface area to radiate heat. DNA evidence also supports an African origin. Modern African populations have more genetic diversity than any other modern human population, inferring that they have been evolving longer (Becoming Human c2008).
Figure \(\PageIndex{26}\): Homo sapiens.
The oldest Homo sapiens fossils were found in Africa at Omo Kibish, Ethiopia, dated to almost 200,000 years ago (195 kya; kya = 1,000 years ago). Homo sapiens fossils found at Herto, Ethiopia (160 kya–154 kya), Klasies River Mouth and Border Cave, South Africa (120 kya – 89 kya), Skhūl, Israel (130 kya–100 kya), and Qafzeh, Israel (120 kya–92 kya), add further support to the age of Homo sapiens. Sites in Asia, e.g., in China and Borneo, Indonesia date to 40 kya or younger and in Europe date no earlier then 31 kya (Mladeč, Czech Republic). Modern humans somehow made it to Australia over open ocean by 55 kya; however, no human remains have been found on the Australian continent dating earlier than 30 kya at Lake Mungo. This data lends little support for the Multiregional Hypothesis, a model that proposed that modern H. sapiens evolved in separate places in the Old World from local archaic H. sapiens populations. This model suggested that gene flow between the populations led to the genetic similarity of modern humans. Most anthropologists support the model of an African origin. There are two variants of this model. The Out of Africa model claims that modern humans arose in Africa and spread to Europe after 50 kya, replacing non-H. sapiens population with no gene flow. An alternative is the Assimilation model that also claimed modern human first arose in Africa and spread to Europe and Asia. The primary difference between the Out of Africa and Assimilation models is that the latter does claim gene flow between H. sapiens and H. neanderthalensis. Taking into account the recent DNA analyses of Neanderthals discussed above, multiple lines of evidence support the Assimilation model.
The case of the disappearing Neanderthals
We know that some Neanderthal genes are in the genome of modern humans, so in essence, some part of the Neanderthals survives today. However, there are no Neanderthals walking around in the modern world. So, what happened to the Neanderthals? Scientists suggest that H. sapiens out-competed Neanderthals with their more diverse diet and “sophisticated and cognitive abilities” (Becoming Human c2008), which may have included superior ability to form visual imagery in imagination to forecast outcomes of future actions, thereby enhancing planning and inventiveness (Koenigshofer, 2017). These traits allowed H. sapiens to readily adapt to rapidly changing climatic conditions during the Upper Paleolithic.
A recent hypothesis suggests that Neanderthals died out during a period of powerful volcanic activity in western Eurasia. Excavations at Mezmaiskaya Cave in the Caucasus Mountains in southern Russia have recovered a plethora of Neanderthal bones, stone tools, and remains of prey animals. The Neanderthal remains and artifacts appear in strata above a layer of volcanic ash and below a second layer of volcanic ash. No Neanderthal bones or artifacts have been found above the second level, suggesting that Neanderthals no longer occupied the area. Both of the ash layers contain levels of pollen associated with cooler, drier climates. “The ash layers correspond chronologically to what is known as the Campanian Ignimbrite super-eruption, which occurred around 40,000 years ago in modern day Italy, and a smaller eruption thought to have occurred around the same time in the Caucasus Mountains” (University of Chicago Press Journals 2010). The ensuing volcanic winter caused a dramatic climate shift that led to the demise of the Neanderthals. This data correlates with Higham et al.’s (2014) research that claims Neanderthals went extinct between 41,000 and 39,000 years ago.
Brain Evolution
Many scientists (Cajal, 1937; Crick & Koch, 1990; Edelman, 2004) believe that the human brain is the most complex machine in the known universe. Its complexity points to one undeniable fact—that it has evolved slowly over time from simpler forms. Evolution of the brain is intriguing not only because we can marvel at this complicated biological structure, but also because it reveals a long history of many less complex nervous systems (Figure 3.3.25), suggesting a record of adaptive behaviors in life forms other than humans. Thus, evolutionary study of the nervous system is important, and it is the first step in understanding its design, its workings, and its functional interface with the environment.
Figure \(\PageIndex{27}\): The brains of various mammals (not to scale). The brains shown are artificially depicted as approximately equal in size. Though the vast differences in the actual size of these brains is not shown, it is still possible to see the increasing degree of folding of the cerebral cortex as indicated by the greater number of fissures and gyri (valleys and hills on the surface of the cortex, indicated by the number of lines) on the surface of the chimpanzee and human brains compared to the others. Note that the mouse cortex is smooth, showing no folding at all indicating less corticalization--cortical development. The greatest folding of the cerebral cortex occurs in humans revealing a trend in mammalian and primate evolution toward increasing corticalization along with increasing brain size. For more on trends in brain evolution in mammals and primates, see section 18.13.
The brains of some animals, other mammals, like apes, monkeys, and rodents, are structurally similar to humans (Figure 3.3.27), while others are not (e.g., invertebrates such as the octopus). Does anatomical similarity of these brains suggest that behaviors that emerge in these species are also similar? Indeed, many animals display behaviors that are similar to humans, e.g., apes use nonverbal communication signals with their hands and arms that resemble nonverbal forms of communication in humans (Gardner & Gardner, 1969; Goodall, 1986; Knapp & Hall, 2009). If we study very simple physiological responses made by individual neurons (nerve cells), the neural responses of invertebrates such as the sea slug (Kandel & Schwartz, 1982) look very similar to the neural responses of single neurons in mammals, including humans, suggesting that from time immemorial such basic functions of individual nerve cells in the brains of many simple animal forms have been conserved over the course of evolution of more complex animals and that in fact these fundamental features of nerve cells are the foundation of more complex behaviors in animals that evolved later (Bullock, 1984).
Although basic physiological processes in nerve cells are very similar across species, nevertheless even at the micro-anatomical level, we note that individual neurons differ in complexity in some respects across animal species. Human neurons exhibit more intricate complexity than neurons in other animals; for example, neuronal processes (dendrites, which receive input from other neurons) in humans have many more branch points, branches, and dendritic spines associated with connections between neurons (see Chapter 10 on Learning and Memory).
Complexity in the structure of the nervous system, both at the macro- and micro-levels, gives rise to complex behaviors. We can observe similar movements of the limbs, as in nonverbal communication, in apes and humans, but the variety and intricacy of nonverbal behaviors using hands in humans surpasses apes. This is related to the fact that humans have more neurons for the control of movement of the hands than apes do. Deaf individuals who use American Sign Language (ASL) express themselves in English nonverbally; they use this language with such fine gradation that many accents of ASL exist (Walker, 1987). Complexity of behavior with increasing complexity of the nervous system, especially the cerebral cortex, can be observed in the genus Homo (Figures 3.3.27 and 3.3.28). If we compare sophistication of material culture in Homo habilis (2 million years ago; brain volume ~650 cm3) and Homo sapiens (300,000 years to now; brain volume ~1400 cm3), the evidence shows that Homo habilis used crude stone tools in stark contrast with modern tools used by Homo sapiens to erect cities, develop written languages, create complex industries, and embark on space travel. All of this is due to increasing complexity of the brain.
Figure \(\PageIndex{28}\): Increasing brain volume and other anatomical and behavioral features including development of material culture (including stone tools) over homininevolution. Note that both brain volume and cultural advances accelerate after first use of fire. Missing from this diagram is the 1,400 cubic centimeter brain of Neanderthals, equal or slightly larger than the brain of modern humans.
What led to the complexity of the brain and nervous system during evolution, and the accompanying refinement of behavior and cognition? Darwin (1859, 1871) proposed two forces: natural selection and sexual selection as work engines driving these changes. As noted previously, Darwin prophesied, “psychology will be based on a new foundation, that of the necessary acquirement of each mental power and capacity by gradation.” In other words, Darwin predicted that psychology will be based on evolution (Rosenzweig, Breedlove, & Leiman, 2002).
Figure \(\PageIndex{29}\): Hominin timeline highlighting evolutionary splits (divergences) from gorilla and chimpanzee genetic lines, major behavioral/cultural innovations, and migration out of Africa. Not shown is migration of Homo sapiens into Europe about 45,000 years ago where they co-existed with Neanderthals for thousands of years (Image from Wikimedia Commons; File:HumanTimeline-TemplateImage-20181222.png; https://commons.wikimedia.org/wiki/F...e-20181222.png; by Drbogdan; licensed under the Creative CommonsAttribution-Share Alike 4.0 International license).
Tool Use, Evolution of Intelligence, and the Brain
As the discussion above attests, tool construction and use is ancient in the genus Homo dating back millions of years at least to Australopithecus. Cachel (2012, p. 222) states, "Human tool behavior is species-specific. It remains a diagnostic feature of humans." According to Osiurak and Massen (2014, p.1107),"Tool use has also been often viewed as an important step during evolution (van Schaik et al., 1999) or even as a marker of the evolution of human intelligence." Vaesen (2012, p. 203) argues that human tool making and tool use provide evidence of "unique higher cognitive ability" in humans and mark "a major cognitive discontinuity" between humans and apes. According to Vaesen (p. 203), there are "nine cognitive capacities deemed crucial to tool use: enhanced hand-eye coordination, body schema plasticity, causal reasoning, function representation, executive control, social learning, teaching, social intelligence, and language. Since striking differences between humans and great apes stand firm in eight out of nine of these domains, [he] conclude[s] that human tool use still marks a major cognitive discontinuity between us and our closest relatives." He also "address[es] the evolution of human technologies. In particular, [he] show[s] how the cognitive traits reviewed help to explain why technological accumulation evolved so markedly in humans, and so modestly in apes." Technological accumulation refers to the accumulation of technological innovations over generations leading to ever increasing technological complexity. This is closely tied to the great capacity of humans for efficient cultural transmission of learned behaviors over successive generations, an ability that occurs in apes only to a relatively limited degree. Vaesen (p. 213) explains: "The complexity of human technologies is tightly linked to our remarkable ability for cumulative culture [bold added]: Humans have been able to build complex systems by accumulating modifications over successive generations, gradually improving on previous achievements." This requires "sophisticated mechanisms for social learning and active teaching," in which language certainly played a part, but it is "likely that other cognitive capacities need to be added; for example, . . . causal thought is part and parcel of human social learning, and hence," is also needed for "high-fidelity cultural transmission" (p. 214). Vaesen further emphasizes this point: "causal thought (whenever it emerged) is a plausible explanans for why cumulative culture evolved so markedly in humans and so modestly in apes" (p.214). Cachel (2012, p. 222) goes further and states that experiments demonstrate the "great gulf that exists between human causal reasoning and the reasoning abilities of chimpanzees. In fact, chimpanzees do not seem to have a “theory of how the world works.” If this is so, then it epitomizes the unique character of human tool behavior – that it is based on the unconscious, sophisticated knowledge of energy, movement, objects, and the interaction of objects that Povinelli et al. (2000) label 'folk physics'.” Causal reasoning lowers individual learning costs, making learning by individuals more efficient, "by making individual learning reasoned rather than random" (Vaesen, 2012, p.215). Vaesen also argues that executive control involving inhibition of current drives and focus of attention contributes in a similar way.
Vaesen (2012) describes the work of other researchers who have identified brain areas involved in tool use. Vaesen (p.204) writes "Orban and colleagues (2006) identified a set of functional regions in the dorsal intraparietal sulcus (IPS) of the human brain that is involved in representations of the central visual field and in the extraction of three-dimensional form from motion. Crucially, these brain regions were not found in the brains of monkeys. The regions subserve, the authors conjectured, the enhanced visual analysis necessary for the precision with which humans manipulate tools. Second, Stout and Chaminade (2007) found that parts of these regions were indeed recruited when modern human subjects engaged in Oldowan-like tool making. Importantly, no increased activation was observed when the human subjects were asked just to strike cobbles together without intending to produce flakes. Human dorsal IPS, thus, may allow for better identification of suitable targets on the core, and as such, explain in part why humans outperform other primates in matters of tool use."
Additional insights regarding tool use and the brain come from study of brain-damaged individuals. Osiurak and Massen (2014, p. 1107) review results of studies of "patients with tool use disorders, also called apraxia of tool use. When asked to light a candle, for example, those patients can light the candle correctly but then put it to the mouth in an attempt to smoke it. Such observations have led traditional cognitive models of apraxia to assume that tool use is supported by sensorimotor knowledge about tool manipulation . . . This sensory-motor hypothesis assumes that manipulation knowledge stored within inferior fronto-parietal areas [of the cerebral cortex] is critical to tool use skills." This may be the case because "the left inferior parietal lobe might rather support technical reasoning, namely, the ability to reason about physical object properties. . . . the ability to understand mechanical actions might be the specificity of the anterior portions of the inferior parietal lobe (particularly the supramarginal gyrus) while the posterior parietal cortex might be involved in the planning of the grasping and reaching components of both tool-use and non-tool-use actions."
Figure \(\PageIndex{30}\): The human right cerebral cortex highlighting parts of the parietal lobe. Folds on the surface of the cortex form hills (gyri, singular, gyrus) and valleys (fissures or sulci; singular, sulcus). The central sulcus divides the frontal lobe from the parietal lobe behind it. The parietal lobe, especially on the right side of the brain, processes information about space and spatial relations. Regions of the parietal lobe are involved in tool construction and tool use, important in human evolution. See text for details. (Image from Wikimedia Commons; File:ParietCapts lateral.png; https://commons.wikimedia.org/wiki/F...ts_lateral.png; author, Sebastian023; licensed under the Creative CommonsAttribution-Share Alike 3.0 Unported license. Caption by Kenneth A. Koenigshofer, PhD).
Tool construction and use, although critically important in human evolution as indicated by the discussion above, are just a single narrow aspect of the complex of abilities that make up human intelligence. In Chapter 14 on Intelligence and Cognition, we continue our discussion of this topic by examining the brain mechanisms which underlie other cognitive abilities that we collectively refer to as intelligence. In the next section of this chapter, David Buss from the University of Texas examines evolutionary theories in psychology.
References
Alemseged Z, Spoor F, Kimbel WH, Bobe R, Geraads D, Reed D, Wynn JG. (2006). A juvenile early hominin skeleton from Dikika, Ethiopoa. Nature 443 (Sep 21): 296-301. Available from: http://www.nature.com.offcampus.lib....ture05047.html. doi:10.1038/nature05047.
Anghelinu, M. (2013). Fitting the ladder to the tree: A common sense view on the cognitive evolution of the Pleistocene human lineage. Arheologia Moldovei, 36 (1), 7-24.
Bailey SE. (2006). Sahelanthropus tchadensis. In: Encyclopedia of anthropology, Vol. 5. Thousand Oaks (CA): SAGE Reference. p. 2044-2045.
Callaway E. (2015). Neanderthals had outsize effect on human biology. Nature [Internet] [cited 2015 Aug 21]; 523 (7562): 512-513. Available from: http://www.nature.com.offcampus.lib....iology-1.18086. doi: 10.1038/523512a
Cooper A, Stringer CB. (2013). Did the Denisovans cross Wallace’s Line? Science [Internet] [cited 2015 Aug 21}; 342 (6156): 321-323. Available from: http://www.sciencemag.org.offcampus....d-963a99b700ea. doi: 10.1126/science.1244869.
Cosmides, L., & Tooby, J. (2002). Unraveling the enigma of human intelligence: Evolutionary psychology and the multimodular mind. The evolution of intelligence, 145-198.
Dembo M, Matzke NJ, Mooers AØ, Collard M. (2015). Bayesian analysis of a morphological supermatrix sheds light on controversial hominin relationships. Proc R Soc B [Internet] [cited 2015 Aug 20]; 282(1812). Available from: http://rspb.royalsocietypublishing.o...0943.e-letters. doi: 10.1098/rspb.2015.0943.
Domínguez-Rodrigo M, Pickering TR, Baquedano E, Mabulla A, Mark DF, Musiba C, et al. (2013). First partial skeleton of a 1.34-million-year-old Paranthropus boisei from Bed II, Olduvai Gorge, Tanzania. PLoS ONE 8(12): e80347. Available from: http://journals.plos.org/plosone/art...l.pone.0080347. doi:10.1371/journal.pone.0080347
Dunbar, R. I. (1998). The social brain hypothesis. Evolutionary Anthropology: Issues, News, and Reviews: Issues, News, and Reviews, 6 (5), 178-190.
eFossils [Internet] [cited 2015 Aug 10]. Department of Anthropology, The University of Texas at Austin. Available from: http://efossils.org/
eLucy: Step-by-step: the evolution of bipedalism [Internet]. c2007. Austin (TX): Department of Anthropology, University of Texas-Austin. [cited 2015 Aug 3]. Available from: elucy.org/Main/LessonOverview.html
Geertz, A. W. (2013). The meaningful brain: Clifford Geertz and the cognitive science of culture. In: Mental Culture: Classical Social Theory and the Cognitive Science of Religion. Edited byDimitris Xygalatas and William W. McCorkle Jr., 176-196.
Gibbons, A. (2012). Generation Gaps Suggest Ancient Human-Ape Split. Science, August 13, 2012. American Association for the Advancement of Science. https://www.science.org/content/arti...%20years%20ago. Retrieved July 4, 2022.
Harmand S, Lewis JE, Feibel CS, Lepre CJ, Prat S, Lenoble A, Boës X, Quinn RL, Brenet M, Arroyo A, Taylor N, Clément S, Daver G, Brugal JP, Leakey L, Mortlock RA, Wright JD, Lokorodi S, Kirwa C, Kent DV, Roche H. 2015. 3.3-million-year-old tools from Lomekwi 3, West Turkana, Kenya. Nature 521 (May 21): 310-315. Available from: http://www.nature.com.offcampus.lib....ture14464.html. doi:10.1038/nature14464.
Higham T, Douka K, Wood R et al. (2014). The timing and spatiotemporal patterning of Neanderthal disappearance. Nature [Internet] [cited 2015 Aug 21]; 512 (7514): 306-309. Available from: http://www.nature.com.offcampus.lib....ture13621.html. doi:10.1038/nature13621
Jurmain R, Kilgore L, Trevathan W. (2013). Essentials of physical anthropology, 9th edition. Belmont (CA): Wadsworth Cengage Learning.
Koenigshofer, K. A. (2017). General Intelligence: Adaptation to Evolutionarily Familiar Abstract Relational Invariants, Not to Environmental or Evolutionary Novelty. The Journal of Mind and Behavior, 38 (2), 119-153.
Larsen, CS. (2014). Our origins: discovering physical anthropology. New York (NY): W. W. Norton & Company, Inc.
Lebatard AE, Bouriès DL, Duriner P, Jolivet M, Braucjer R, Carcaillet J, Schuster M, Arnaud N, Monié P, Lihoreau F, Likius A, Macaye HT, Vignaud P, Brunet M. 2008. Cosmogenic nuclide dating of Sahelanthropus tchadensis and Australopithecus bahrelghazali: Mio-Pliecene hominids from Chad. Proc Natl Acad Sci U S A [Internet] [cited 2015 Aug 3]; 105(9): 3226-3231. Available from: http://www.pnas.org/content/105/9/3226.full. doi: 10.1073/pnas.0708015105
Lice and human evolution [Internet]. 2011 Feb 6. NOVAscienceNow. [cited 2015 Aug 20]. Available from: www.pbs.org/wgbh/nova/evolution/lice.html
Meyer M, Kircher M, Gansauge MT, et al. (2012). A high-coverage genome sequence from an archaic Denisovan individual. Science [Internet] [cited 2015 Aug 21]; 338 (6104): 222-226. Available from: http://www.sciencemag.org.offcampus..../6104/222.full. doi: 10.1126/science.1224344.
O’Neil D. c1999-2012. Early human evolution: a survey of the biological and cultural evolution of Homo habilis and Homo erectus. Behavioral Sciences Department, Palomar College [Internet] [cited 2015 Aug 16]. Available from: anthro.palomar.edu/homo/Default.htm
O’Neil D. c1999-2014. Evolution of modern humans: a survey of the biological and cultural evolution of archaic and modern Homo sapiens. Behavioral Sciences Department, Palomar College [Internet] [cited 2015 Aug 16]. Available from: anthro.palomar.edu/homo2/default.htm
Osiurak, F., & Massen, C. (2014). The cognitive and neural bases of human tool use. Frontiers in psychology, 5, 1107.
Oxnard, C., Obendorf, P.J., Kefford, B.J. (2010). Post-cranial skeletons of hyperthyroid cretins show a similar anatomical mosaic as Homo floesiensis. PLoS One [Internet] [cited 2015 Aug 21]; 5 (9). Available from: http://www.sciencedaily.com/releases...0928025514.htm. doi: 10.1371/journal.pone.0013018.
Reich D, Green RE, Kircher M, et al. (2010). Genetic history of an archaic hominin group from Denisova Cave in Siberia. Nature [Internet] [cited 2015 Aug 21]; 468 (7327). Available from: http://www.nature.com/nature/journal...ture09710.html
Richmond BG, Jungers WL. (2008). Orrorin tugenensis femoral morphology and the evolution of hominin bipedalism. Science [Internet] [cited 2015 Aug 3]; 319(5870): 1662-1665. Available from: www.jstor.org/stable/20053635
Scarre C. (2005).The Human Past: World Prehistory&the Development of Human Societies. London (UK): Thames & Hudson Ltd.
Scarre C. (2013). The human past. London (UK): Thames & Hudson.
Su DF. (2013). The earliest hominins: Sahelanthropus, Orrorin, and Ardipithecus. The Nature Education Knowledge Project [Internet] [cited 2015 Aug 3]. Available from: http://www.nature.com/scitable/knowl...hecus-67648286
Vaesen, K. (2012). The cognitive bases of human tool use. Behavioral and brain sciences, 35 (4), 203-218.
Ward CV, Manthi FK, Plavcan JM. (2013). New fossils of Australopithecus anamensis from Kanapoi, West Turkana, Kenya (2003-2008). J Hum Evol 65(5): 501-524. Available from doi:10.1016/j.jhevol.2013.05.006
Wood, B. (2010). Reconstructing human evolution: Achievements, challenges, and opportunities. Proceedings of the National Academy of Sciences, 107 (supplement_2), 8902-8909.
University of Chicago Press Journals [Internet]. Volcanoes wiped out Neanderthals, new study suggests. ScienceDaily 2010 Oct 7 [cited 2015 Aug 21]. Available from: http://www.sciencedaily.com/releases...1006094057.htm
Bullock, T. H. (1984). Comparative neuroscience holds promise for quiet revolutions. Science, 225(4661), 473–478.
Cachel, S. (2012). Human tool behavior is species-specific and remains unique. Behavioral and Brain Sciences, 35 (4), 222.
Crick, F., & Koch, C. (1990). Towards a neurobiological theory of consciousness. Seminars in the Neurosciences, 2, 263–275.
Darwin, C. (1871). The descent of man, and the selection in relation to sex. London: J. Murray.
Darwin, C. (1859). On the origins of species by means of natural selection, or, The preservation of favoured races in the struggle for life. London, UK: J. Murray.
Demonet, J. F., Chollet, F., Ramsay, S., Cardebat, D., Nespoulous, J. L., Wise, R., . . . Frackowiak, R. (1992). The anatomy of phonological and semantic processing in normal subjects. Brain, 115(6), 1753–1768.
Edelman, G. (2004). Wider than the sky: The phenomenal gift of consciousness. New Haven, CT: Yale University Press.
Gardner, R. A., & Gardner, B. T. (1969). Teaching sign language to a chimpanzee. Science, 165(3894), 664–672.
Goodall, J. (1986). The chimpanzees of Gombe: Patterns of behavior. Cambridge, MA: Harvard University Press.
Hubel, D. H. (1995). Eye, brain, and vision. Freeman & Co., NY: Scientific American Library/Scientific American Books.
Knapp, M. L., & Hall, J. A. (2009). Nonverbal communication in human interaction. Boston, MA: Wadsworth Cengage Learning.
Osiurak, F., & Massen, C. (2014). The cognitive and neural bases of human tool use. Frontiers in psychology, 5, 1107.
Ramón y Cahal, S. (1937). Recollections of my life. Memoirs of the American Philosophical Society, 8, 1901–1917.
Rolls, E. T., & Cowey, A. (1970). Topography of the retina and striate cortex and its relationship to visual acuity in rhesus monkeys and squirrel monkeys. Experimental Brain Research, 10(3), 298–310.
Rosenzweig, M. R., Breedlove, S. M., & Leiman, A. L. (2002). Biological psychology (3rd ed.). Sunderland, MA: Sinauer Associates.
Smith, E. E., & Jonides, J. (1999). Storage and executive processes in the frontal lobes. Science, 283(5408), 1657–1661.
Smith, E. E., & Jonides, J. (1997). Working memory: A view from neuroimaging. Cognitive psychology, 33, 5–42.
Van Essen, D. C., Anderson, C. H., & Felleman, D. J. (1992). Information processing in the primate visual system: An integrated systems perspective. Science, 255(5043), 419–423.
Walker, L. A. (1987). A loss for words: The story of deafness in a family. New York, NY: Harper Perennial.
Attributions
"Human Evolution" text and figures not attributed to other sources adapted by Kenneth A. Koenigshofer, PhD, Chaffey College, from Biological Anthropology (Saneda and Field), chapter: Human Evolution; LibreTexts by Tori Saneda & Michelle Field, Professors (Anthropology) at Cascadia Community College (via Wikieducator)
"Defining Hominins" text and figures not attributed to other sources were adapted and modified by Kenneth A. Koenigshofer, Ph.D., Chaffey College, from Explorations: An Open Invitation to Biological Anthropology; https://explorations.americananthro.....php/chapters/; Chapter 9: "Early Hominins," Editors: Beth Shook, Katie Nelson, Kelsie Aguilera and Lara Braff, American Anthropological Association Arlington, VA 2019; CC BY-NC 4.0 International, except where otherwise noted.
"Homo Genus," "Defining the Genus Homo," "Climate," "Homo habilis: The Earliest Members of Our Genus," "Homo erectus," "Homo erectus Culture," "Tool Use and Cognitive Abilities of Homo erectus" and "The Big Picture of Early Homo," text and figures not attributed to other sources adapted by Kenneth A. Koenigshofer, PhD, Chaffey College, from Explorations: An Open Invitation to Biological Anthropology; https://explorations.americananthro.....php/chapters/; Chapter 10, "Early Members of the Genus Homo" by Bonnie Yoshida-Levine Ph.D.; Beth Shook, Katie Nelson, Kelsie Aguilera, and Lara Braff, Eds.; licensed under a Creative Commons Attribution-NonCommercial 4.0 International License, except where otherwise noted.


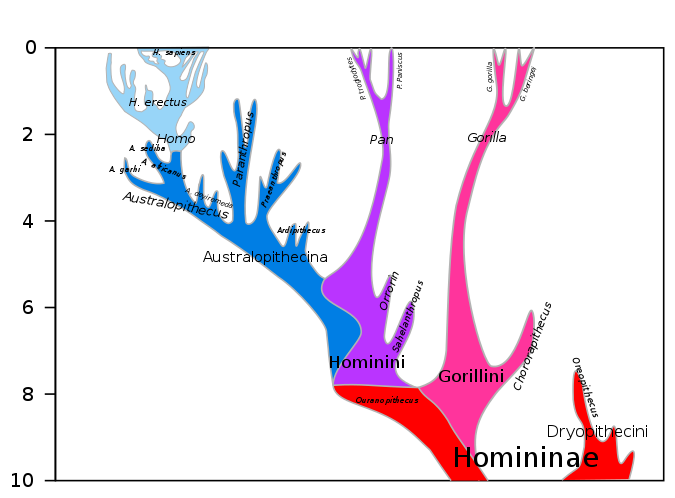
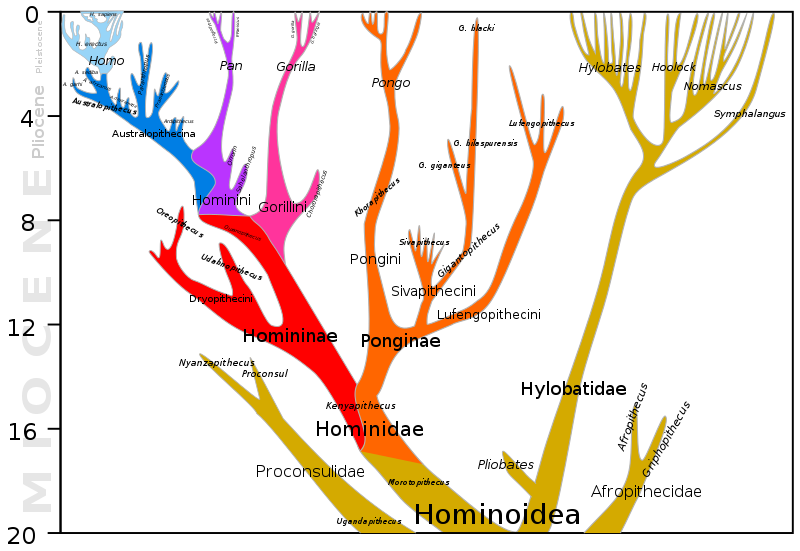


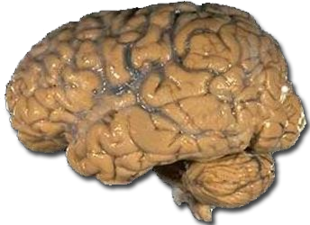
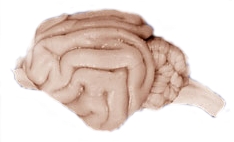

.jpg?revision=1&size=bestfit&width=398&height=400)
.jpg?revision=1&size=bestfit&width=348&height=349)

.jpg?revision=1&size=bestfit&width=310&height=413)
.jpg?revision=1)