
5.1: Neurons and their Basic Functions
- Page ID
- 110437
This page is a draft and under active development. Please forward any questions, comments, and/or feedback to the ASCCC OERI (oeri@asccc.org).
Learning Objectives
- Describe the anatomy of neurons and the function of each of the three major parts of a neuron
- Describe neurotransmitters, synapse, synaptic vesicles, resting potential, EPSP, IPSP, and action potential
- Explain saltatory conduction and why it is important to communication within the nervous system; include description of the myelin sheath and nodes of Ranvier
- Describe the role of post-synaptic receptor sites in communication between neurons
- Describe neurons classified by shape and by their functions
Overview
In this section, we continue our exploration of neurons, the building blocks of the nervous system. We examine how they generate electrochemical signals, and how the billions of neurons in the nervous system communicate with one another, a process known as synaptic transmission. Before tackling these topics, we review and expand the basic anatomy and functioning of neurons covered in part in Chapter 4. A sound grasp of these facts provides the groundwork for understanding how neuron potentials are generated within neurons and how they combine to trigger synaptic transmission. As you read, remember that the voltages and chemical events we discuss in this section, operating in large populations of brain cells, somehow generate our perceptions, thoughts, emotions, and the entirety of our mental experience. To date, how this happens, how patterns of neuron potentials in brain circuits become conscious minds, remains the greatest mystery of all facing modern science.
Introduction to Neuron Anatomy, Neuron Potentials, and Synaptic Transmission
The nervous system of vertebrates like us is comprised of the central nervous system, made up of the brain and spinal cord, and the peripheral nervous system, which consists of all the nerves outside the brain and spinal cord.
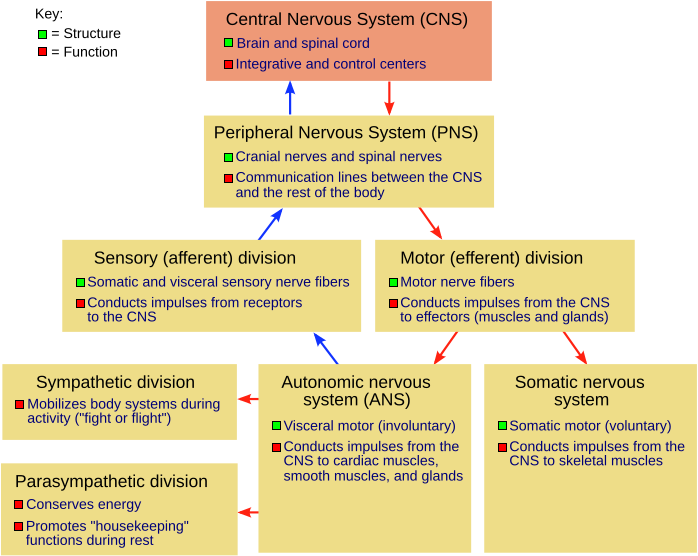
Figure \(\PageIndex{1}\): The human nervous system and its major divisions. All components consist of neurons--nerve cells. (NSdiagram NGB colors fixed.png adapted by Naomi Bahm (colors boxed and fixed) from File:NSdiagram.svg; https://commons.wikimedia.org/wiki/File:NSdiagram.svg; by Fuzzform via Wikimedia Commons; Permission is granted to copy, distribute and/or modify this document under the terms of the GNU Free Documentation License, Version 1.2 or any later version published by the Free Software Foundation; with no Invariant Sections, no Front-Cover Texts, and no Back-Cover Texts. A copy of the license is included in the section entitled GNU Free Documentation License. Subject to disclaimers).
The peripheral nervous system is composed of the somatic sensory nerves (input nerves such as the auditory nerve, the optic nerve, spinal sensory nerves mediating skin sensations, etc.) and somatic motor nerves (output nerves, serving the skeletal muscles, activating or inhibiting them) as well as the autonomic sensory and motor nerves (serving the internal body organs such as the heart, lungs, blood vessels, digestive system, adrenal glands, etc.). These autonomic motor nerves, which either activate or inhibit the internal body organs, are of two types: the sympathetic (activate heart, lungs, constrict blood vessels, etc. to prepare the body for "fight or flight") and parasympathetic (conserve the body's resources during non-emergency, non-threatening situations) motor autonomic nerves. All of these parts of the nervous system, including the brain, spinal cord, and the peripheral nervous system, are made up of nerve cells, technically known as neurons. The human brain is estimated to have somewhere between 80 and 100 billion neurons. Recall from Chapter 4 that neurons have three major parts: the soma or cell body, the dendrites which receive inputs from other neurons, and the axon, which carries the output of neurons, nerve impulses (action potentials), to other neurons, to muscles, or to glands.
You have probably seen photographs or drawings of neurons previously in your introductory psychology course. The images below show small sections of cerebral cortex in a human adult and a human infant. Note the density of neurons and connections in each brain slice. The complexity of interconnections among neurons is evident in the image at the far right which is based on use of Golgi stain.
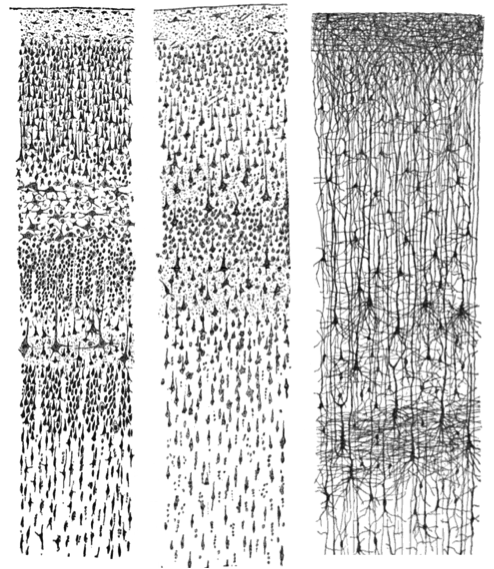
Figure \(\PageIndex{2}\): Small sections of tissue from the brain of a human infant and a human adult. The image at the far right using Golgi stain highlights cell bodies and extensive branching of dendrites. Left: Nissl-stained visual cortex of a human adult. Middle: Nissl-stained motor cortex of a human adult. Right: Golgi-stained cortex of a 1 1/2 month old infant. [Image: Santiago Ramon y Cajal, https://goo.gl/zOb2l1, CC0 Public Domain, https://goo.gl/m25gce]
Nissl and Golgi Stains
Note that different chemicals were used to stain the neurons in the figure above so that they could be seen under the microscope. Nissl stain labels only the main part of the cell (i.e., the cell body; see left and middle panels of Figure 5.1.2). However, by contrast, a Golgi stain fills the cell body and all the processes that extend outward from the cell body (see right panel of Figure 5.1.2). A more notable characteristic of a Golgi stain is that it only stains approximately 1–2% of neurons (Pasternak & Woolsey, 1975; Smit & Colon, 1969), permitting the observer to distinguish one cell from another. These qualities of the Golgi stain allowed the pioneering neuroanatomist Santiago Ramon y Cajal to examine the full anatomical structure of individual neurons for the first time. This significantly enhanced our appreciation of the intricate networks their processes form. Based on his observation of Golgi-stained tissue, Cajal suggested neurons were distinguishable processing units rather than part of a continuous network of nerves, as had been previously believed. Cajal and Golgi shared the Nobel Prize for Medicine in 1906 for their combined contribution to the advancement of science and our understanding of the structure of the nervous system.
Neuron Anatomy and Basic Functions
To understand neuron function, it is important to be familiar with the anatomy involved. As noted above, neurons have three major structural parts - - the soma or cell body, the axon (carries the neuron's output), and the dendrites (the "receivers" of the neuron). The entire neuron is bounded by a cell membrane, the neural membrane. The cell membrane of a neuron has channels or "doors" for ions (electrically charged atoms) which can pass through the membrane when specific channels are opened for specific ions. Figure 5.1.4 shows this basic neuron anatomy. Various types of neurons are discussed below.
The soma or cell body contains organelles, common to all types of cells in the body. These are involved in the basic metabolism of the cell. The soma also contains the nucleus, where the genes and chromosomes (containing DNA) are located.
The second main part of the neuron is the dendrites, the receivers of the neuron. Dendrites in some neurons can branch profusely (large numbers of dendritic branches off the main shaft of a dendrite with their own branches are often collectively called dendritic trees), expanding the region of the neuron that can receive inputs from other neurons. The receptor sites (or more technically, postsynaptic receptor sites because of their location on the receiving or postsynaptic neuron) which receive molecules of neurotransmitter are located on the dendrites (and, to a lesser degree, on the soma) of the receiving neuron. On the dendrites are small dendritic spines which are associated with the connections between neurons (the synapses) and can change shape rapidly when learning occurs (see Chapter 10 for more detail). Note that the spines are not the same thing as dendritic branches. In Figure 5.1.3, a dendritic branch with dendritic spines is shown on the left in microscopic detail, and on the right, are dendritic trees of two types of neuron found in the retina. Spines are not visible in the images of dendritic trees on the right because the dendritic spines are too small, while dendritic branches comprising the dendritic trees can easily be seen (see caption for Figure 5.1.3).

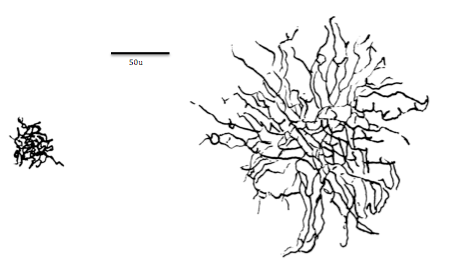
Figure \(\PageIndex{3}\): (Left) A segment of pyramidal cell dendrite from stratum radiatum (CA1) of the hippocampus with thin, stubby, and mushroom-shaped spines. Spine synapses are colored in red, stem (or shaft) synapses are colored in blue. The dendrite was made transparent in the lower image to enable visualization of all synapses. (Right) A size comparison between midget and parasol cell dendritic trees. Dendritic spines are too small to be visible in the two images on the right. Parasol and midget neurons are found in the ganglion cell layer of the retina. (On left, Pyramidal cell dendrite and spines: Image and caption from Wikimedia Commons; File:A segment of pyramidal cell dendrite from stratum radiatum (CA1).jpg; https://commons.wikimedia.org/wiki/F...atum_(CA1).jpg; by Synapse Web, Kristen M. Harris, PI, http://synapses.clm.utexas.edu/; licensed under Creative Commons Attribution 2.0 Generic license. On right, Midget and parasol cell dendritic trees: Image and one line caption from Wikimedia Commons; File:Midget vs Parasol cell.png; https://commons.wikimedia.org/wiki/F...rasol_cell.png; by Stromdabomb; licensed under the Creative Commons Attribution-Share Alike 4.0 International license; two sentence explanation by Kenneth A. Koenigshofer, PhD).
The third major part of the neuron is the axon, coming out of the soma like a hose. The axon carries the output messages of a neuron (nerve impulses) along its length to its axon terminal buttons (axon endings). There is only one axon per neuron, although it can branch into multiple axon terminal buttons. In a typical neuron, the root end of the axon emerges out of the soma at a small swelling called the axon hillock. Between the axon hillock and the first segment of the axon is where the nerve impulse is first generated (see discussion of the action potential, the nerve impulse, that follows below).

Figure \(\PageIndex{4}\): Basic structure of a neuron (Image from Wikimedia Commons; File:Components of neuron.jpg; https://commons.wikimedia.org/wiki/F..._of_neuron.jpg; by Jennifer Walinga; licensed under the Creative Commons Attribution-Share Alike 4.0 International license).
Myelinated Axons and Saltatory Conduction
Some axons have a glial cell covering (glials are non-neural cells in the nervous system, see Chapter 4.1) known as the myelin sheath. This fatty, insulating, myelin sheath has gaps in it, revealing the bare axon, at regular intervals along the axon's length. These bare spots along the length of a myelinated axon are called nodes, or nodes of Ranvier, after their discoverer.
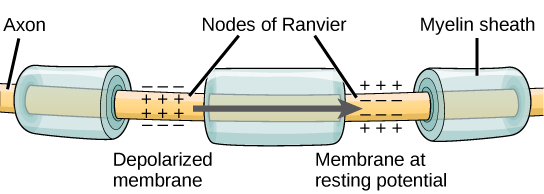
Figure \(\PageIndex{5}\): Nodes of Ranvier. Nodes of Ranvier are gaps in the myelin sheath which covers myelinated axons. Nodes contain voltage-gated potassium (K+) and sodium (Na+) channels. Action potentials travel down the axon by "jumping"from one node to the next, speeding conduction of the action potential down the length of the axon toward the axon ending, also known as the axon bouton, axon button, or axon terminal.
The function of the myelin sheath and the nodes is to speed up the rate at which nerve impulses travel down the length of the axon toward their destination, the axon ending (axon bouton). In myelinated axons, the impulses sort of "jump" from node to node allowing the action potential to move more rapidly down the axon. This leaping of the nerve impulse (action potential) from node to node is called saltatory conduction, from the Latin "saltatore" which means to dance. Imagine the romantic image of the impulse dancing from node to node.
A node of Ranvier is a natural gap in the myelin sheath along the axon. These unmyelinated spaces are about one micrometer long and contain voltage-gated (i.e., opened by voltage) sodium (Na+) and potassium (K+) ion channels (ions are electrically charged atoms). The flow of ions through these channels, particularly the Na+ channels, regenerates the action potential over and over again along the axon at each successive node of Ranvier. As noted above, the action potential “jumps” from one node to the next in saltatory conduction. If nodes of Ranvier are not present along an axon (as is the case in unmyelinated axons; see below), the action potential propagates much more slowly; ion movement through Na+ and K+ channels continuously regenerates new action potentials at every successive point along the axon, using extra time to do so. In effect, conduction of the action potential along an axon involves voltage-gated channels (channels that respond to voltage, rather than to neurotransmitter), on the axon, responding to voltage arising from an electrical field which spreads from the action potential in the previous segment of the axon. In a myelinated axon, because the action potential jumps from node to node, it does not have to be regenerated at every successive point along the axon, but only at the nodes, skipping across myelinated segments of the axon between nodes. Because the action potential in a myelinated axon must be regenerated fewer times to move any particular distance along the length of the axon, it reaches its destination faster, compared to the speed of conduction in an unmyelinated axon.
Nodes of Ranvier also save energy for the neuron since the ion channels only need to be present and opened and closed at the nodes and not along the entire axon. It is extraordinary that the nodes are placed along the axon's length at just the right spatial intervals to make impulse conduction down the axon the most efficient and speedy as possible. One can only wonder at the incredible precision with which natural selection operated on this feature of myelinated axons over the long course of animal evolution to create this optimal spacing of the nodes.
Neural Conduction in Unmyelinated Axons
Not all axons are myelinated. Unmyelinated axons tend to be older in evolution and to be the smaller diameter axons (classified as C fibers based on their small diameters; large diameter myelinated axons are called A fibers). In unmyelinated axons, in order to move, the nerve impulse must be regenerated at every successive point along the axon. This takes time and slows the conduction of the nerve impulse (the action potential) down the length of the axon. Therefore, conduction of the action potential down the length of an unmyelinated axon is relatively slow. An example of unmyelinated C fibers are axons that are part of slow pain pathways. These pathways mediate the slower aching pain that follows tissue damage. The quick, sharp pain from an injury is mediated by larger diameter A fibers (axons).
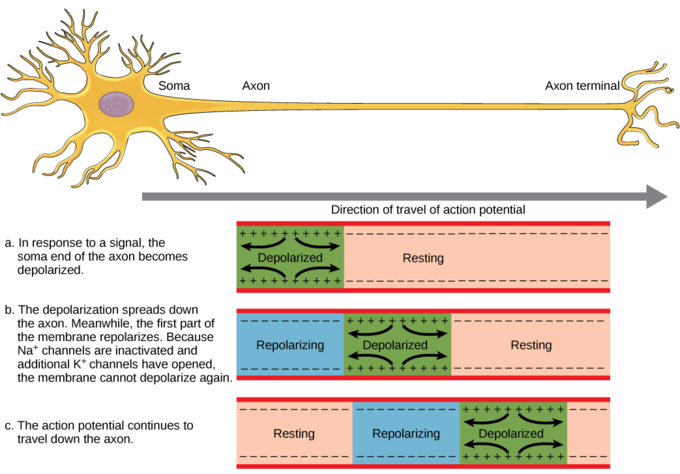
Figure \(\PageIndex{6}\): Action potential traveling along an unmyelinated neuronal axon. The action potential is conducted down the axon as the axon membrane depolarizes, then repolarizes. Because of these dynamics, the action potential can only be conducted in one direction, away from the cell body. Image: figure-35-02-04_NGB_added_resting.png adapted by Naomi Bahm (resting added to part a) from https://s3-us-west-2.amazonaws.com/c...e-35-02-04.png
The ion channels along the length of the unmyelinated axon are called voltage-gated channels because the voltage generated by the action potential in the prior segment of the axon triggers opening of these channels in the next segment. This leads to regeneration of the action potential. As this process is repeated over and over along the length of the unmyelinated axon, the action potential is conducted along the length of the axon away from the cell body toward the axon endings of the neuron. Figure \(\PageIndex{7}\) shows the channels opening and closing and the ions moving across the cell membrane generating an action potential in successive segments of the axon. This causes movement of the action potential, its conduction, along the axon away from the cell body (from right to left in the figure, as the arrow shows).
Figure \(\PageIndex{7}\): Another depiction of propagation of the nerve impulse (the axon potential) along an axon. This animation illustrates action potential propagation in an axon. Three types of ion channel are shown: potassium "leak" channels (blue), voltage-gated sodium channels (red) and voltage-gated potassium channels (green). The movement of positively-charged sodium and potassium ions through these ion channels controls the membrane potential of the axon. Action potentials are initiated in the axon's initial segment after neurotransmitter activates excitatory receptors in the neuron's dendrites and cell body. This depolarizes the axon initial segment to the threshold voltage for opening of voltage-gated sodium channels. Sodium ions entering through the sodium channels shift the membrane potential to positive-inside. The positive-inside voltage during the action potential in the initial segment causes the adjacent part of the axon membrane to reach threshold voltage. When positive-inside membrane potentials are reached, voltage-gated potassium channels open and voltage-gate sodium channels close. Potassium ions leaving the axon through voltage-gated potassium channels return the membrane potential to negative-inside values. When the voltage-gated potassium channels gate shut, the membrane potential returns to the resting potential. (Image and caption from Wkimedia Commons; File:Action potential propagation animation.gif; https://commons.wikimedia.org/wiki/F..._animation.gif; by John Schmidt; licensed under the Creative Commons Attribution-Share Alike 4.0 International, 3.0 Unported, 2.5 Generic, 2.0 Generic and 1.0 Generic license).
By contrast, as described above, in myelinated axons, the impulse gets regenerated only at the bare spots on the axon, the nodes of Ranvier. Because the action potential gets regenerated fewer times in order to travel a given distance, than is the case for unmyelinated axons, neural conduction is faster with the insulating myelin sheath. Myelinated axons tend to be large diameter (A and B fibers) and found in neural pathways mediating rapid behavioral response, such as the Pyramidal tract that runs from motor cortex to spinal cord motor neurons which generate voluntary action.
Synapses
The axon splits many times, so that it can communicate, or synapse, with several other neurons (see Figure 5.1.4). At the end of the axon, the axon terminal button (also called the axon ending, the terminal bouton, or the axon terminal) forms synapses with the dendritic spines, small protrusions mentioned above, on the dendrites of receiving neurons (postsynaptic neurons). Synapses form between the axon terminal button of the presynaptic neuron (neuron sending the signal) and the postsynaptic membrane (membrane of the neuron receiving the signal; see Figure 5.1.4). Here we will focus specifically on synapses between the axon terminal button of an axon and a dendritic spine; however, synapses can also form between the axon terminal button of the presynaptic neuron and the postsynaptic neuron's soma, dendritic shaft directly, or the axon of another neuron.
A very small space called a synaptic gap or a synaptic cleft exists between the pre-synaptic axon's terminal button and the post-synaptic neuron's dendritic spine. To give you an idea of the size of the synaptic gap, a dime is 1.35 mm (millimeter) thick. There are 1,350,000 nm (nanometers) in the thickness of a dime. The synaptic gap is only about 5 nm wide. In the pre-synaptic terminal button, there are synaptic vesicles that package together groups of chemicals, neurotransmitters (see Figure 5.1.4). Neurotransmitters are released from the pre-synaptic axon's terminal button or axon ending into the synaptic gap; molecules of neurotransmitter then travel across the synaptic gap, and open ion channels on the post-synaptic spine by binding to receptor sites there. We will discuss the role of these receptors in more detail later in section 5.2.
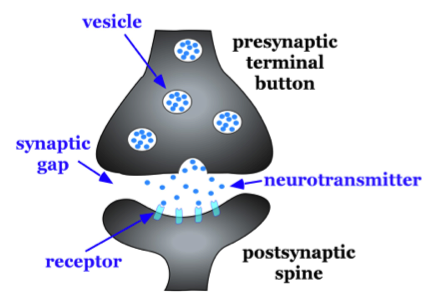
Figure \(\PageIndex{8}\): Basic characteristics of a typical synapse. Enlargement of the synapse between one of the axon terminal buttons (labeled presynaptic terminal button) and one of the dendrites of the second neuron shown (on the right) in Figure 5.1.4 above.
Axon Terminal Buttons, Synaptic Vesicles, PSPs, and Synaptic Transmission
Recall that the end of an axon or axon branch is called the axon terminal button (or terminal, or simply the axon ending). Within the axon terminal button are structures called synaptic vesicles, which contain neurotransmitter chemicals (which have been manufactured in the soma and transported to the axon ending and stored there in the synaptic vesicles, ready for release). When neurons communicate with one another across the synaptic gap which separates them, it is the neurotransmitter, released from the synaptic vesicles in the axon terminal button of the "sender" neuron (the presynaptic neuron) that transmits the neural message across the synaptic gap (the space between the membrane of the axon terminal button of the pre-synaptic neuron and the membrane of the dendrite or soma of the postsynaptic neuron). The event that triggers release of neurotransmitter from the synaptic vesicles in the axon terminal button is the arrival of an action potential at the axon terminal button of the sender neuron. The nerve cell releasing the neurotransmitter, the sender neuron, is known technically as the presynaptic neuron. The neuron receiving the neurotransmitter, the receiver cell, is called the postsynaptic neuron.
Figure \(\PageIndex{9}\): A synapse. Synaptic vesicles release neurotransmitters (small yellow balls) which bind to the receptors (blue peg-like structures) on the postsynaptic membrane. Synaptic vesicles inside a presynaptic axon terminal button (axon ending) releasing neurotransmitter molecules onto receptors on a dendrite of a receiving (post-synaptic) neuron. (Image from Wikimedia Commons; File:Neurotransmitters.jpg; https://commons.wikimedia.org/wiki/F...ansmitters.jpg; by https://www.scientificanimations.com/; by https://www.scientificanimations.com/; licensed under the Creative Commons Attribution-Share Alike 4.0 International license).
The neurotransmitter molecules attach to special sites on the membrane of the postsynaptic neuron. These special sites are called receptor sites or postsynaptic receptor sites and are typically located on the dendrites and dendritic spines of the receiving neuron, but can also be on its soma. Their molecular shapes match the shapes of the neurotransmitter molecules they receive, a kind of "lock and key" fit, which opens chemically-gated ion channels allowing ions with their electrical charges to move across the cell membrane creating voltage shifts called postsynaptic potentials (PSPs; additional details follow in Section 5.2).
These events from release of transmitter to generation of post-synaptic potentials comprise synaptic transmission. Many psychoactive drugs (drugs that alter mind and/or behavior) such as amphetamines, LSD, "magic mushrooms," etc. produce their effects by blocking or activating specific receptor sites (other psychoactives produce their effects by other mechanisms which affect synaptic transmission, some of which are discussed below and in Chapter 6 of this textbook).
As mentioned above, the cell membrane of a neuron has channels or "doors" for ions (electrically charged atoms) which can pass through the membrane when specific channels are opened for specific ions. Normally the channels are closed until acted upon by the attachment of neurotransmitter molecules to receptors sites on the membrane of the receiving neuron (the post-synaptic neuron). The attachment of the molecules of transmitter to the receptors sites, like a key into a lock, opens the "doors" (ion channels) to specific ions which pass through the cell membrane carrying with them their electrical charge. This results in a change in the electrical state of the neuron, a post-synaptic potential (i.e. a voltage shift in the post-synaptic neuron)--if a positive shift in voltage occurs, it is called an EPSP, an excitatory post-synaptic potential; if a negative shift in voltage occurs in the post-synaptic neuron, then it is called an IPSP, an inhibitory post-synaptic potential. EPSPs are caused by excitatory neurotransmitters (e.g. glutamate; acetylcholine; histamine; epinephrine), IPSPs by inhibitory transmitters (e.g. GABA, gamma amino butyric acid; serotonin, also known by its chemical name, 5-hydroxytryptamine, abbreviated 5-HT).
These PSPs, whether excitatory or inhibitory, are called graded potentials because they are not of a fixed voltage, but instead vary in voltage depending on the amount of neurotransmitter (and other factors) that has been released onto the receptor sites on the postsynaptic neuron's dendrite. This is in contrast to the action potential (the nerve impulse) which is of a fixed voltage and is "all or none" which means that if it occurs, it occurs at its full strength or not at all. Think of a gun firing. If you pull the trigger of the gun with sufficient force (a kind of "trigger threshold" of force), then the gun fires a bullet with its full strength. But if trigger threshold is not reached, the gun doesn't fire at all. You don't get half a shot or one-eighth of a shot if you pull with less force, you can only get an "all or none" result--the gun fires, or it doesn't. Either the force on the trigger reaches threshold and the gun fires at its full "strength," or if the force is insufficient, the gun does not fire a bullet. The same is true of the action potential; if an action potential occurs, it occurs "all or none," at its full strength (voltage), or not at all, and, like the gun, the neuron "fires" only if its trigger threshold or firing threshold (in voltage, about -55 millivolts for most neurons) is reached. Graded potentials (EPSPs and IPSPs) are akin to analog signals (of different voltages), whereas action potentials aresimilar to digital signals (fixed voltage).
Types of Neurons
Not all neurons are created equal! There are neurons that receive information about the world around us, sensory neurons. There are motor neurons that allow us to initiate movement and behavior, ultimately allowing us to interact with the world around us. Finally, there are interneurons, which process the sensory input from our environment into meaningful representations, plan the appropriate behavioral response, and connect to the motor neurons to execute these behavioral plans.
Figure \(\PageIndex{10}\): Sensory neurons carry information towards the CNS. Motor neurons carry information from the CNS. Interneurons carry information between sensory and motor neurons.
In addition to this general functional categorization of neurons (sensory, motor, and interneurons), neurons may also be classified by structure (e.g. unipolar, bipolar, multipolar), shape, or other characteristics. See Chapter 4.1 for an overview of these classifications. Since the shape of the neuron relates to its role in communication, this categorization is explored in greater depth below.
Categorizing Neurons by Shape
Neurons can also be classified by their shape. For example, Cajal used such names as basket, stellate, moss, pyramidal, etc. to describe various types of neurons he observed. However, this classification is anatomical and does not reflect whether the cell is, for example, a motor neuron or an interneuron. Nevertheless, the shape of neurons may reflect aspects of their function and their role in information processing in the nervous system. For example, pyramidal neurons (Figure \(\PageIndex{11}\)) are "a common class of neuron found in the cerebral cortex of virtually every mammal, as well as in birds, fish and reptiles. Pyramidal neurons are also common in subcortical structures such as the hippocampus and the amygdala. They are named for their shape: typically they have a soma (cell body) that is shaped like a teardrop or rounded pyramid. They also tend to have a conical spray of longer dendrites that emerge from the pointy end of the soma (apical dendrites) and a cluster of shorter dendrites that emerge from the rounded end. . . They comprise about two-thirds of all neurons in the mammalian cerebral cortex . . . they are ‘projection neurons’ — they often send their axons for long distances, sometimes out of the brain altogether. For example, pyramidal neurons in layer 5 of the motor cortex send their axons down the spinal cord to drive muscles" (Bekkers, 2011). They are the primary excitatory neurons in the motor cortex and the pre-frontal cortex. Table \(\PageIndex{1}\) summarizes features of pyramidal neurons.
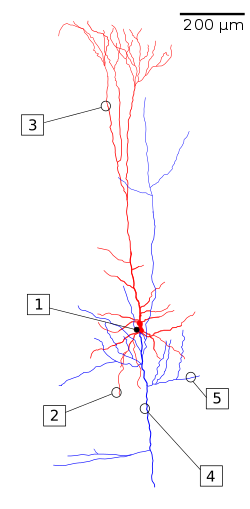 Figure \(\PageIndex{11}\): (left) A reconstruction of a pyramidal cell. Soma and dendrites are labeled in red, axon arbor in blue. (1) Soma, (2) Basal dendrite, (3) Apical dendrite, (4) Axon, (5) Collateral axon. (right) Pyramidal neuron of a rat hippocampal organic culture. Axons shown (axones, in Spanish) are from another neuron most of which is not shown (Image on left, Wikipedia, The Pyramidal Cell, retrieved 8/30/21. Image on right from Wikimedia Commons, File:Neurone pyramidal.jpg; https://commons.wikimedia.org/wiki/F..._pyramidal.jpg; by Mathias De Roo; licensed under the Creative Commons Attribution-Share Alike 3.0 Unported license.). |
|
| Table \(\PageIndex{1}\). Features of pyramidal neurons. (adapted from John Bekkars; Pyramindal Neurons; Current Biology). | |
|---|---|
| Location | Common in cerebral cortex of mammals, especially Layers III and V (cerebral cortex in mammals has six cell layers); also found in hippocampus, and amygdala. Also found in other vertebrates |
| Shape and numbers | Multipolar Pyramidal, but some variations among species and location in brain; about 2/3 of all neurons in the mammalian cerebral cortex |
| Function | Excitatory projection neurons with long axons carrying action potentials long distances, sometimes completely out of the brain; e.g. pyramidal neurons in layer 5 of motor cortex go all the way to spinal motor neurons; others that remain in the cortex interconnecting distant areas of cortex are critically important in many cognitive functions |
| Neurotransmitter | Glutamate, the most abundant excitatory transmitter in the vertebrate nervous system |
By contrast, stellate neurons have a star-like shape by virtue of the dendrites which radiate out from the stellate neuron's cell body (see Figure 5.1.12(b)). Spiny stellate neurons, like pyramidal neurons, have large numbers of dendritic spines, but they lack the long apical dendrite characteristic of pyramidal neurons. Like pyramidal neurons, spiny stellate neurons in the cerebral cortex are excitatory and use glutamate as transmitter. Van Essen and Kelly (1973) reported different functions for visual cortical neurons depending upon whether they were pyramidal or stellate neurons. After pyramidal neurons, stellate neurons are the second most numerous type of cortical neuron. Another type of stellate neuron that has only sparse spines is inhibitory and uses GABA (gamma-amino-butyric-acid) as its transmitter.
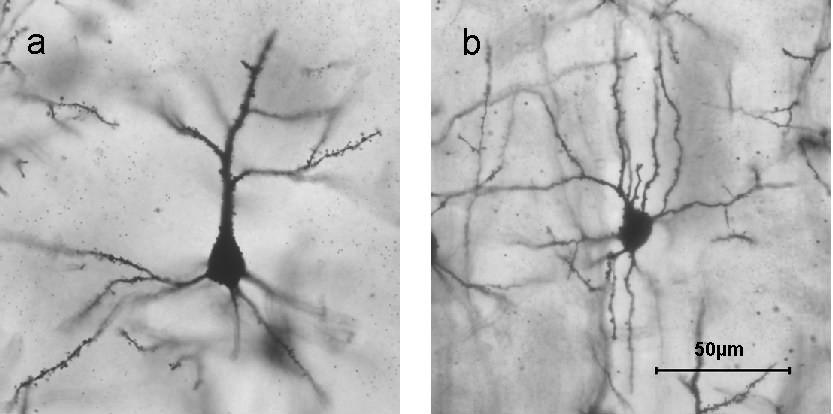
Figure \(\PageIndex{12}\): Golgi stained neurons in different layers of cerebral cortex: a) Layer II/III pyramidal cell; b) layer IV spiny stellate cell. Dendritic spines are visible on branching dendrites as clusters of tiny thorn-like bristles. (Image and one sentence caption from Wikipedia, Stellate Cell, retrieved 8/30/21; description of dendritic spines by Kenneth A. Koenigshofer, Ph.D.).
Brown et al. (2019) found different processing roles for basket neurons (see Figure 5.1.13) and stellate neurons in the cerebellum. Stellate cells and basket cells both make inhibitory synapses directly onto Purkinje neurons in the cerebellum. Stellate neurons influence pattern of firing and basket neurons affect rate of firing of the Purkinje neurons in the cerebellum. Basket cells are also found in the neocortex (cerebral cortex). Basket cells are inhibitory interneurons. They envelop pyramidal cell bodies in the neocortex in dense complexes resembling baskets (Kirkcaldie, 2012). Figure \(\PageIndex{13}\) shows a variety of interneurons classified by shape.
"Approximately 95% of the cortical neuronal activity is mediated by fast excitatory (glutamate, 80%) and fast inhibitory (GABA, 15%) neurons. The remaining 5% percent is associated with the slow modulatory action of monoaminergic (dopamine, serotonin, noradrenaline) and non-monoaminergic (acetylcholine, endorphins, etc.) neurons located in small subcortical nuclei of the mesencephalon and projecting to the cerebral cortex" (Marco Catani, in Encyclopedia of Behavioral Neuroscience, 2nd edition, 2022, Science Direct; https://www.sciencedirect.com/topics...pyramidal-cell; retrieved 4/25/22).
Though several basic types of neurons have been classified, the picture in the brain with regard to types of neurons present is quite complex, as expressed in this quote: "Whereas in the spinal cord we could easily distinguish neurons based on their function [sensory, interneuron, motor], that isn’t the case in the brain. Certainly, there are brain neurons involved in sensory processing – like those in visual or auditory cortex – and others involved in motor processing – like those in the cerebellum or motor cortex. However, within any of these sensory or motor regions, there are tens or even hundreds of different types of neurons. In fact, researchers are still trying to devise a way to neatly classify the huge variety of neurons that exist in the brain . . . part of what gives the brain its complexity is the huge number of specialized neuron types. Researchers are still trying to agree on what these are" (University of Queensland, n.d.).
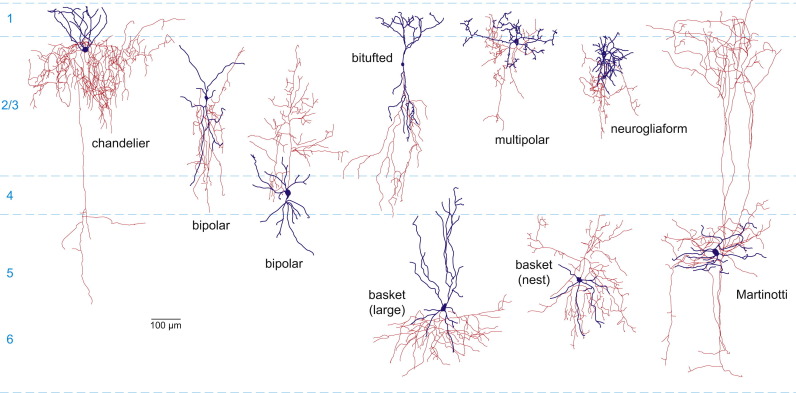
Figure \(\PageIndex{13}\): (above). Variety of types of interneurons classified by shape. Representative morphologies of mouse interneuron types. Note that the axon (red) and dendrite (blue) arbors are typically less elaborate than in primate cortex. Two variants of bipolar cells and basket cells (large and nest) are shown (Kirkcaldie, 2012).
Yet another way to classify neurons is by the neurotransmitter they use (see Synaptic Transmission below). Perhaps equally important is to attempt to trace the connections that neurons make with other neurons to try to understand their inputs and outputs and what roles they play in processing. This has been done for many stellate and basket cells, inhibitory interneurons synapsing on Purkinje cells in the cerebellum. However, because of the complexity of brain circuitry this is very difficult to accomplish, especially in the cerebral cortex, although possible in cases where neurons belong to large neural tracts or pathways with straightforward connections which are easier to trace, such as from the pyramidal neurons in motor cortex to spinal cord motor neurons or from thalamus to sensory cortex. Understanding the complex circuitry of the brain in its entirety may never be fully accomplished. With 80-100 billion neurons in the human brain and most making perhaps many thousands of connections, the number of connections in the human brain is astronomical!
References
Bekkers, J. M. (2011). Pyramidal neurons. Current biology, 21(24), R975.
Brown, A. M., Arancillo, M., Lin, T., Catt, D. R., Zhou, J., Lackey, E. P., ... & Sillitoe, R. V. (2019). Molecular layer interneurons shape the spike activity of cerebellar Purkinje cells. Scientific reports, 9 (1), 1-19.
Churchland, P. M. (2013). Matter and consciousness. MIT press.
Fields, R. D., & Stevens-Graham, B. (2002). New insights into neuron-glia communication. Science, 298 (5593), 556-562.
Kirkcaldie, M. T. (2012). Neocortex. In The mouse nervous system (pp. 52-111). Academic Press.
Koenigshofer, K.A. (2011). Mind Design: The Adaptive Organization of Human Nature, Minds, and Behavior. Pearson Education, Boston.
Mishra A, Singh S, Shukla S. (2018). Physiological and Functional Basis of Dopamine Receptors and Their Role in Neurogenesis: Possible Implication for Parkinson's disease. J Exp Neurosci. 12:1179069518779829. [PMC free article] [PubMed]
Nadim, F., & Bucher, D. (2014). Neuromodulation of neurons and synapses. Current opinion in neurobiology, 29, 48-56.
Pasternak, J. F., & Woolsey, T. A. (1975). On the "selectivity" of the Golgi-Cox method. J Comp Neurol, 160(3), 307-312. doi: 10.1002/cne.901600304
Perea, G., Sur, M., & Araque, A. (2014). Neuron-glia networks: integral gear of brain function. Frontiers in cellular neuroscience, 8, 378.
Smit, G. J., & Colon, E. J. (1969). Quantitative analysis of the cerebral cortex. I. Aselectivity of the Golgi-Cox staining technique. Brain Res, 13(3), 485-510.
University of Queensland, Queensland Brain Institute, Types of Neurons, n.d. https://qbi.uq.edu.au/brain/brain-an.../types-neurons; Retrieved 8/31/21
Van Essen, D., & Kelly, J. (1973). Correlation of cell shape and function in the visual cortex of the cat. Nature, 241(5389), 403-405.
Attributions
1. Chapter 5, Communication within the Nervous System, section 5.1., "Neurons and their Basic Functions," written by Kenneth A. Koenigshofer, Ph.D. (except material listed in attributions below), is licensed under CC BY 4.0
2. Figure 5.1.2, Vocabulary, Discussion Questions, Outside Resources, and "Nissl and Golgi Stains" adapted by Kenneth A. Koenigshofer from: Furtak, S. (2021). Neurons. In R. Biswas-Diener & E. Diener (Eds), Noba textbook series: Psychology. Champaign, IL: DEF publishers. Retrieved from http://noba.to/s678why4
Creative Commons License



 Neurons by Sharon Furtak is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License. Permissions beyond the scope of this license may be available in our Licensing Agreement.
Neurons by Sharon Furtak is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License. Permissions beyond the scope of this license may be available in our Licensing Agreement.
3. Figures 5.1.4., 5.1.5, and some text adapted from: General Biology (Boundless), Chapter 35, The Nervous System;
https://bio.libretexts.org/Bookshelv...gy_(Boundless); LibreTexts content is licensed by CC BY-NC-SA 3.0. Legal.
4. Figures 5.1.9 and 5.1.10 adapted from Chapter 11.3 (Neurons and Glia Cells), 11.4 (Nerve Impulses) in Book: Human Biology (Wakim & Grewal) - Biology LibreTexts by Suzanne Wakim & Mandeep Grewal, under license CC BY-NC
Video Series: Neurobiology/Biopsychology - Tutorial animations of action potentials, resting membrane potentials, and synaptic transmission.
- http://www.sumanasinc.com/webcontent/animations/neurobiology.html
- Video: An animation and an explanation of an action potential
- Video: An animation of neurotransmitter actions at the synapse
- Video: An interactive animation that allows students to observe the results of manipulations to excitatory and inhibitory post-synaptic potentials. Also includes animations and explanations of transmission and neural circuits.
- https://apps.childrenshospital.org/clinical/animation/neuron/
- Video: Another animation of an action potential
- Video: Another animation of neurotransmitter actions at the synapse
- Video: Domino Action Potential: This hands-on activity helps students grasp the complex process of the action potential, as well as become familiar with the characteristics of transmission (e.g., all-or-none response, refractory period).
- Video: For perspective on techniques in neuroscience to look inside the brain
- Video: The Behaving Brain is the third program in the DISCOVERING PSYCHOLOGY series. This program looks at the structure and composition of the human brain: how neurons function, how information is collected and transmitted, and how chemical reactions relate to thought and behavior.
- http://www.learner.org/series/discoveringpsychology/03/e03expand.html
- Video: You can grow new brain cells. Here\\\'s how. -Can we, as adults, grow new neurons? Neuroscientist Sandrine Thuret says that we can, and she offers research and practical advice on how we can help our brains better perform neurogenesis—improving mood, increasing memory formation and preventing the decline associated with aging along the way.
- Web: For more information on the Nobel Prize shared by Ramón y Cajal and Golgi
- http://www.nobelprize.org/nobel_prizes/medicine/laureates/1906/