
10.6: Localization of Types and Stages of Memory
- Page ID
- 139939
This page is a draft and under active development. Please forward any questions, comments, and/or feedback to the ASCCC OERI (oeri@asccc.org).
Learning Objectives
- Explain the role of the dorsolateral prefrontal cortex in memory, problem-solving and reasoning.
- Discuss working memory and its various components including the central executive, the phonological loop, episodic buffer, and visuospatial sketch pad
- Describe the region of brain associated with the operation of the central executive
- Describe the brain structures involved in consolidation of new memories into long-term storage
- Explain the different types of memory, three stages of memory, and the brain areas associated with each as discussed below
- Describe the role of place cells in the formation of spatial maps in the hippocampus
- Explain the trisynaptic circuit of the hippocampus and its role in memory
Overview
Here we examine memory from the standpoint of its different types focusing on the role of the hippocampus and other medial temporal lobe structures and circuits in the consolidation of new declarative memories. The hippocampal/mammillothalamic tract, also called Papez's circuit, plays an essential role in this process. After consolidation, long-term memories are stored in the cerebral cortex and become independent of the hippocampus. We also consider the role of the prefrontal cortex in working memory, important for the retention of information during problem-solving and reasoning. In addition, we discuss the role of the hippocampus in spatial memory and in the formation of cognitive maps or mental models of space.
Types and Stages of Memory Localized in Different Brain Regions
A large body of evidence indicates that the dorsolateral prefrontal cortex plays an important role in certain forms of memory work (Barbey, et al., 2013), in particular those that involve alternating between two memory tasks and exploring various possibilities before making a choice (Hunt et al., 2015). It seems fairly certain that dorsolateral prefrontal cortex holds information required for reasoning processes that are in progress.
Figure \(\PageIndex{1}\): Divisions of Human Prefrontal Cortex (Image from Wikimedia Commons; File:Prefrontal cortex.png; https://commons.wikimedia.org/wiki/F...tal_cortex.png; by Natalie M. Zahr, Ph.D., and Edith V. Sullivan, Ph.D.; image is a work of the National Institutes of Health and is in the public domain).
Short-term Memory
Short-term memory depends on the attention paid to the elements of sensory memory. Short-term memory lets you retain a piece of information for less than a minute and retrieve it during this time. One typical example of its use is the task of repeating a list of items that has just been read to you, in their original order. In general, you can retain 5 to 9 items (or, as it is often put, 7±2 items) in short-term memory. It is clear that short-term memory does not depend on the medial temporal cortex including the hippocampus and surrounding structures. We know this because H.M. who had bilateral removal of his medial temporal lobe still retained his short-term explicit memory. If someone came into the room, he could remember the visit for a short time. Of course, as you have learned, if we checked his recollection a few minutes later, without ability to consolidate the memory into a long-term form, he would have no recollection of the visit that occurred just minutes before. This suggests that the dorsolateral prefrontal cortex, left intact in H.M., may be at least one of the structures capable of retaining information for a brief period, particularly when engaged in reasoning and decision-making. But its precise role remains the subject of much debate.
Working Memory
Working memory is a more recent extension of the concept of short-term memory. As techniques for studying memory have become more refined, it has become increasingly apparent that the original conception of short-term memory as a mere temporary receptacle for long-term memory is too simplistic. In fact, it is becoming increasingly clear that there is no strict line of demarcation between memories and thoughts. In order to test some hypotheses that may provide a better understanding of this complex phenomenon, the concept of working memory has therefore been advanced.
Working memory is used to perform cognitive processes on the items that are temporarily stored in it. It would therefore be heavily involved in processes that require reasoning, such as reading, or writing, or performing computations. One typical example of the use of working memory is the task of repeating a list of items that has just been read to you, but in the reverse of their original order. Another good example is the task of simultaneous language interpretation, where the interpreter must store information in one language while orally translating it into another. In the course of a day, there are many times when you need to keep some piece of information in your head for just a few seconds. Maybe it is a number that you are “carrying over” to do a subtraction, or a persuasive argument that you are going to make as soon as the other person finishes talking. These are good examples of why you usually hold information in your short-term/working memory: to accomplish something that you have planned to do. Perhaps the most extreme example of working memory is a chess master who can explore several possible solutions mentally before choosing the one that will lead to checkmate.
Working memory appears to be composed of several independent systems, which would imply that we are not aware of all the information that is stored in it at any given time. For example, when you drive a car, you are performing several complex tasks simultaneously. It is unlikely that all of the various types of information involved are being handled by a single short-term memory system.
Baddeley & Hitch (1974) proposed a model of working memory with several components: a certain number of auxiliary “slave” systems and a central executive that supervises and controls the flow of information to and from the "slave systems," which are short-term storage repositories for particular types or domains of information. One of these slave systems, the phonological or articulatory loop, specializes in processing linguistic information, while another, the visuospatial sketchpad, specializes in processing visual-spatial information. The episodic buffer is a third storage system, dedicated to linking information across domains to form integrated units of visual, spatial, and verbal information (e.g., the memory of a story, a movie scene, or an integrated autobiographical memory). The episodic buffer is also assumed to have links to long-term memory.
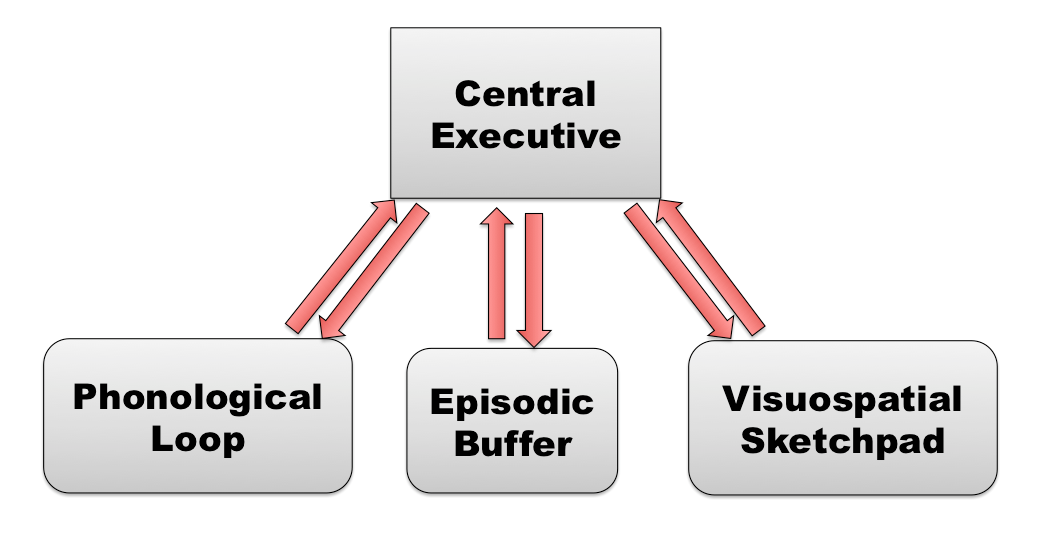
Figure \(\PageIndex{2}\): Baddeley's Model of Working Memory. (Image from Wikimedia Commons; Baddeley's Model of Working Memory; https://commons.wikimedia.org/wiki/F...ing_Memory.png; author, AmandaSilver15; permission is granted to copy, distribute and/or modify this document under the terms of the GNU Free Documentation License, Version 1.2 or any later version published by the Free Software Foundation).
The phonological or articulatory loop plays an important role in everyday life. For example, when you repeat a phone number to yourself in your head, you are activating this loop. This loop is also heavily involved in reading and writing. The presence in working memory of another slave system that can manipulate mental images of visual objects is suggested by tests where subjects are asked to rotate such images.
Perhaps the most important but least understood component in Baddeley’s model of working memory is the central processor or central executive, whose job would be to select, initiate, and halt the routines performed by its slave systems.
Baddeley’s model of working memory has proven especially fruitful for research on the brain areas involved in problem solving. This model posits a central executive or central processor that coordinates the activity of two sub-systems. Many brain-imaging studies show high activity in the frontal lobe, in particular the prefrontal lobe, when this central executive is working. For example, the image shown below (see Figure 10.5.3.2) was produced by functional magnetic resonance imaging (fMRI). As you have learned in a previous chapter, fMRI is a technique based on the increased blood flow to the most active areas of the brain. In this image, taken while the subject was holding an image of a face in his memory, the yellow area in the prefrontal cortex is very active, confirming the role of these prefrontal cortical areas in working memory.
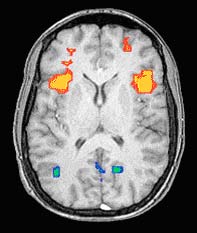
Figure \(\PageIndex{3}\): fMRI images of brain showing increased activity in the frontal lobes while the subject is keeping an image of a face in his memory (Image from NIMH Laboratory of Brain and Cognition. Published in Nature, Vol 386, April 10, 1997, p. 610, Courtney et al., 1997).
As you have learned in an earlier chapter, this region, at the very front of the brain, is highly developed in humans. It appears that the prefrontal cortex, by holding information in storage during working memory, may play a significant role in functions related to attention, problem-solving, and intelligence (Conway et al., 2003; Engle et al., 1999; Kane & Engle, 2002; Matzel and Kolata, 2010; Barbey et al., 2014; Waltz et al., 1999).
Recall that Baddeley’s model also postulates the existence of a phonological (acoustic and linguistic) memory and a visual/spatial memory (containing mental images). Brain imaging studies have also revealed distinct neuroanatomical bases for both of these forms of short-term working memory.
The phonological loop activates certain areas in the left hemisphere that are associated with the production of language, such as Wernicke’s area in the temporal lobe and Broca’s area in the frontal lobe. Visual/spatial memory seems to be associated with a region of the occipital cortex generally associated with visual processing. This is consistent with the hypothesis that visual imagination in which humans can manipulate visual images in the "mind's eye" to mentally test out and anticipate future consequences of action involves parts of the visual system "recruited" into this new function during evolution (Koenigshofer, 2017).
Meanwhile, certain sub-regions of the prefrontal cortex are activated only if the memorization exercise is somewhat difficult for the subject, thus confirming the coordinating role of the central executive.
The phenomenon of working memory is made all the more complex by the fact that it takes place over time. For example, the experimental results illustrated below show how various areas of the subjects’ brains alter their activity levels as the subjects are presented with various visual stimuli. When the subjects are shown a blurred image, the activity level (represented by the blue bars in the graph) becomes highest in area 1, the visual part of the brain. When the subjects are shown an image of a face, brain activity (black bars) becomes highest in the associative and frontal regions (4, 5, and 6). Lastly, when the subjects are retaining an image of a face in their working memory, brain activity (red bars) is highest in the frontal regions, while the visual areas are scarcely stimulated at all.
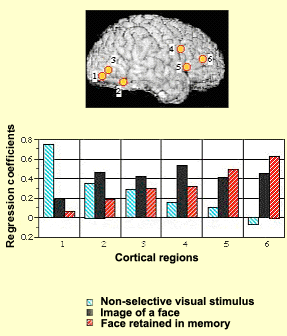
Figure \(\PageIndex{4}\): Activity in cortical areas (labeled 1-6 in image of cortical surface) primarily in occipital lobe with presentation of blurred image (blue bars, tallest in area 1), in frontal and association cortical areas with presentation of a photo of a face (black bars), and in frontal areas (areas 5 and 6), with little activity in visual areas (red bars in cortical areas 1 and 2), when subject retains memory of a face in working memory (red bars). (Image source: NIMH Laboratory of Brain and Cognition. Published in Nature, Vol. 386, April 1997, p. 610, Courtney et al., 1997).
It has also been observed that distinct processes appear to be involved in the storage and recall of items memorized with the phonological loop and the visual/spatial sketchpad. One thing is certain: the prefrontal cortex plays a fundamental role in working memory. It enables people to keep information available that they need for their current reasoning processes. For this purpose, the prefrontal cortex must cooperate with other parts of the cortex from which it extracts information for brief periods. For this information to eventually pass into longer-term memory, the limbic system has to be brought into play.
Long-Term Memory
Recent research has provided a complex, highly intricate picture of memory functions and their loci in the brain. The hippocampus, the temporal lobes, and the structures of the limbic system that are connected to them are essential for the consolidation of long-term declarative memory. The hippocampus receives connections from the cortex’s primary sensory areas, unimodal associative areas (those that involve only one sensory modality), and multimodal associative areas, as well as from the rhinal and entorhinal cortexes. While these anterograde connections converge at the hippocampus, other, retrograde pathways emerge from it, returning to the primary cortexes, where they record in the cortical synapses the associations facilitated by the hippocampus. Thus, in the mechanism of memorization, we find feedback loops among these various brain structures.
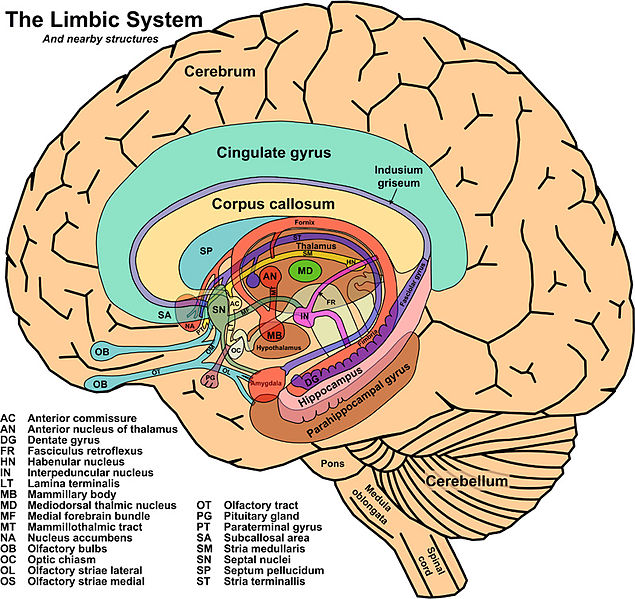
Figure \(\PageIndex{5}\): Limbic system and surrounding structures (Image from Wikimedia; Structures of the Limbic System; https://commons.wikimedia.org/wiki/F...ohn_Taylor.jpg; This work has been released into the public domain by its author, John Taylor).
Hippocampus
The hippocampus facilitates associations among various parts of the cortex, for example, between a tune that you heard at a dinner party represented in auditory cortex in the temporal lobe and the faces of the other guests who were at the table, represented in the visual areas and frontal areas of cortex. However, all other things being equal, such associations would naturally fade over time, so that your mind did not become cluttered with useless memories. What might cause such associations to be strengthened and eventually etched into long-term memory very often depends on “limbic” factors, such as how interested you were in the occasion, or what emotional charge it may have had for you, or how gratifying you found its content.
Recall from prior modules of this chapter that the hippocampus, the cortical structures surrounding it, and the neural pathways that connect them to the cortex as a whole are all heavily involved in declarative memory–the memory of facts and events. Declarative (explicit) memory is composed of semantic memory (memory for facts and knowledge) and episodic memory (memory for episodes in your life).
Semantic memory is the system that you use to store your knowledge of the world. It is a knowledge base that we all have and much of which we can access quickly and effortlessly. It includes our memory of the meanings of words–the kind of memory that lets us recall not only the names of the world’s great capitals, but also social customs, the functions of things, and their color and odor and other sensory properties.
Semantic memory also includes our memory of the rules and concepts that let us construct a mental representation of the world without any immediate perceptions. Its content is thus abstract and relational and is associated with the meaning of verbal symbols.
The hippocampus also plays a fundamental role in episodic memory (sometimes called autobiographical memory), the kind of memory that lets you remember events that you personally experienced at a specific time and place. For example, episodic memory lets you remember an especially pleasant dinner party years later. In fact, it seems to be the hippocampus that enables you to “play the scene back”, by reactivating this particular activity pattern in the various regions of the cortex. This phenomenon would be very important during dreams, and would explain the incorporation of events from the last few days into them.
The various structures of the limbic system exert their influence on the hippocampus and the temporal lobe via Papez’s circuit, also known as the hippocampal/mammillothalamic tract. This circuit is a sub-set of the numerous connections that the limbic structures have with one another. The route that information travels in this circuit is from the hippocampus to the mammillary bodies of the hypothalamus, then on to the anterior thalamic nucleus, the cingulate cortex, and the entorhinal cortex, before finally returning to the hippocampus (see Figure 10.5.5).
Once the temporary associations of cortical neurons generated by a particular event have made a certain number of such “passes” through Papez’s circuit, they will have undergone a physical remodeling that consolidates them. Eventually, these associations will have been strengthened so much that they will stabilize and become independent of the hippocampus. As you have already learned, bilateral lesions of the hippocampus will prevent new long-term memories from forming, but will not erase those that were encoded and consolidated before the injury.
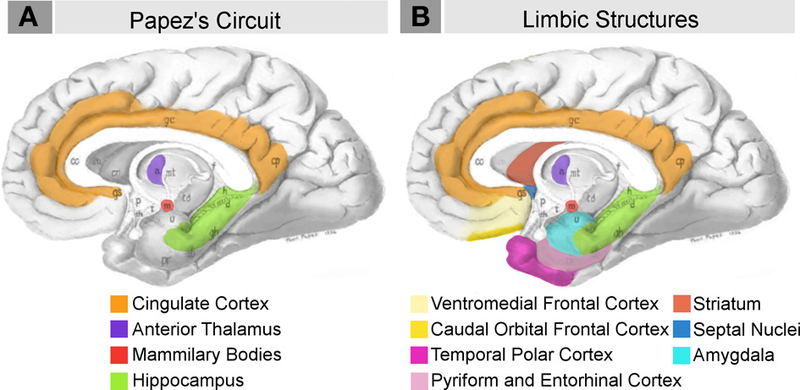
Figure \(\PageIndex{6}\): Papez Circuit and Limbic system structures some of which are involved in memory and emotion. (Images from Wikimedia Commons; File:Neural systems proposed to process emotion.png; https://commons.wikimedia.org/wiki/F...ss_emotion.png; by Barger N, Hanson KL, Teffer K, Schenker-Ahmed NM and Semendeferi K; licensed under the Creative Commons Attribution 3.0 Unported license. Caption by Kenneth A. Koenigshofer, PhD).
With this gradual disengagement of the limbic system, the memories will no longer pass through Papez’s circuit, but instead will be encoded in specific areas of the cortex: the same ones where the sensory information that created the memories was initially received (the occipital cortex for visual memories, the temporal cortex for auditory memories, etc.).
Some very intense personal memories that bring what is sometimes called emotional memory into play appear to involve another structure of the limbic system besides the hippocampus. This structure is the amygdala, located beneath temporal lobe cortex near the hippocampus, and not surprisingly, as discussed in an earlier chapter, the amygdala is already known to manage our reactions to fear. Emotional memories can be very strong and particularly long-lasting. The release of norepinephrine during highly emotional events may contribute to the rapid and effective consolidation of memory for emotionally-charged events.
Injury to the Limbic System Pathways and Memory
As described above, for a piece of information to be recorded in long-term memory, it must pass through Papez’s circuit. Injuries to this circuit can result in memory impairments. For example, a lesion in the mammillary bodies is responsible for an amnesic syndrome whose most classic example is Korsakoff’s syndrome. In addition to the confabulation, confusion, and disorientation that accompany this syndrome, patients suffer from anterograde amnesia: they cannot store new information in their long-term memory. The most typical cause of this syndrome is vitamin B1 deficiency, often seen in chronic alcoholics.
Spatial Memory and a Role in Episodic Memory
The hippocampus appears to play a fundamental role in spatial memory in many animal species, including humans. For example, in a British study, the researchers asked taxi drivers to imagine their travels through the city of London, while their brain activity was monitored by positron emission tomography (PET scan). This task, which was so familiar to these subjects, caused a specific activation of their right hippocampus. Unlike our memory of facts and events, spatial memory appears to be confined to the hippocampus. And more specifically to the right hippocampus. This structure seems to be able to create a mental map of space, thanks to certain cells called place cells.
An amazing discovery in the 1970s demonstrated that a rat’s hippocampus is a veritable spatial map of the environment through which it moves. Certain pyramidal neurons in area CA1 of the rat hippocampus become active only when the rat is located in a specific part of its environment. There are 1 million of these “place cells” in area CA1 of the rat hippocampus, so that if each one is assigned a specific point in space, together they can form a very precise cognitive map that tells the animal where it is at any given time. Moreover, when a rat explores a new environment, it forms a new cognitive map that can be very stable, lasting weeks or months.
According to O’Keefe and Nadel (1978), the researchers who discovered the existence of these cognitive maps, one of their functions might be to provide a context to which episodic memories can be attached, in particular, their location in space and time. An event recorded in memory could thus be made autobiographical (situated in personal time and space). This would explain the fundamental role that the hippocampus plays in episodic memory in humans.
The hippocampus proper is composed of regions with tightly packed pyramidal neurons, mainly areas CA1, CA2, and CA3. (“CA” stands for Cornu Ammonis, or Horn of Ammon. The reference is to the ram’s horns of the Egyptian god Ammon, whose shape these three areas together roughly resemble.) This is what is called the trisynaptic circuit or trisynaptic loop of the hippocampus. Information enters this one-way loop via the axons of the entorhinal cortex (part of the medial temporal cortex), known as perforant fibers (or the perforant path, because it penetrates through the subiculum and the space that separates it from the dentate gyrus, which is also in the temporal lobe, adjacent to the hippocampus; some anatomists consider it part of the hippocampus). These axons of the perforant path make the loop’s first connection, with the granule cells of the dentate gyrus, the first region where all sensory modalities merge together to form representations and memories that bind stimuli together (Science Direct; https://www.sciencedirect.com/topics...0and%20memory.), a critical step in learning and memory.
From granule cells of the dentate, the mossy fibres in turn project to make the loop’s second connection, with the dendrites of the pyramidal cells in area CA3 of the hippocampus.
The axons of these CA3 cells divide into two branches. One branch forms the commissural fibers that project to the contralateral hippocampus via the corpus callosum. The other branch forms the Schaffer collateral pathways that make the third connection in the loop, with the cells in area CA1 of hippocampus. It is in these synapses that the spatial memory, associated with the hippocampus, seems to be encoded.
This region also displays a high propensity for long-term potentiation (LTP), though this same phenomenon is also observed in many other parts of the hippocampus as well as in the cortex.
Lastly, the axons of the cells in CA1 project to the neurons of the subiculum and of the entorhinal cortex. The receiving portion of the hippocampal formation thus consists of the dentate gyrus, while the sending portion consists of the subiculum. The axons of the large pyramidal neurons of the subiculum then project to the subcortical nuclei via the fimbria, a thin tract of white matter at the inner edge of the hippocampus (see Figure 10.4.8). Lastly, the information returns to the sensory cortical areas from which it came before it was processed by the hippocampus.
Thus, entorhinal cortex projects strongly to dentate gyrus and hippocampus and then from hippocampus back to cortex (Van Hoesen and Pandya, 1975b). Being situated between cortex and the hippocampus, entorhinal cortex plays a critical role in memory (Zola-Morgan et al., 1989). Even though the neurons of the hippocampus may seem like just a transit point in the establishment of long-term memory, they actually display a great deal of plasticity. This plasticity, as you have learned in previous sections, is manifested chiefly through long-term potentiation (LTP), which was first discovered in the hippocampus but has subsequently been demonstrated in many regions of the cortex.
Summary
Memory can be understood in terms of types and stages. Different brain structures and circuits are involved in these different types and stages of memory. Dorsolateral prefrontal cortex appears to hold information required for reasoning and decision making processes. The prefrontal cortex is important for retention of information during working memory. Working memory has several components which interact and which have links to long-term memory. Consolidation of new declarative memories depends upon feedback loops between the cortex, the hippocampus, and other limbic structures in the medial temporal lobes. A group of these structures involved in memory, called Papez's circuit or the hippocampal/mammillothalamic tract, includes the hippocampus, cingulate cortex, mammillary bodies, and anterior thalamus. Damage to the mammillary bodies, commonly due to B1 vitamin deficiency associated with alcoholism, causes Korsakoff's amnesia. The hippocampus in rats forms spatial cognitive maps involving the use of hippocampal pyramidal neurons that function as place cells which code particular locations in the environment that the rat moves through.
References
Baddeley, A. D. (2000). The episodic buffer: a new component of working memory? Trends in Cognitive Science, 4, 417-423.
Baddeley, A.D., & Hitch, G. (1974). Working memory. In G.H. Bower (Ed.), The psychology of learning and motivation: Advances in research and theory (Vol. 8, pp. 47--89). New York: Academic Press.
Barbey, A. K., Colom, R., Paul, E. J., & Grafman, J. (2014). Architecture of fluid intelligence and working memory revealed by lesion mapping. Brain Structure and Function, 219 (2), 485-494.
Barbey, A. K., Koenigs, M., & Grafman, J. (2013). Dorsolateral prefrontal contributions to human working memory. cortex, 49 (5), 1195-1205.
Conway, A. R., Kane, M. J., & Engle, R. W. (2003). Working memory capacity and its relation to general intelligence. Trends in cognitive sciences, 7(12), 547-552.
Courtney, S. M., Ungerleider, L. G., Keil, K., & Haxby, J. V. (1997). Transient and sustained activity in a distributed neural system for human working memory. Nature, 386 (6625), 608-611.
Engle, R. W., Kane, M. J., & Tuholski, S. W. (1999). Individual differences in working memory capacity and what they tell us about controlled attention, general fluid intelligence, and functions of the prefrontal cortex. Models of working memory: Mechanisms of active maintenance and executive control, 4, 102-134.
Hunt, L. T., Behrens, T. E., Hosokawa, T., Wallis, J. D., & Kennerley, S. W. (2015). Capturing the temporal evolution of choice across prefrontal cortex. Elife, 4, e11945.
Kane, M. J., & Engle, R. W. (2002). The role of prefrontal cortex in working-memory capacity, executive attention, and general fluid intelligence: An individual-differences perspective. Psychonomic bulletin & review, 9(4), 637-671.
Koenigshofer, K. A. (2017). General Intelligence: Adaptation to Evolutionarily Familiar Abstract Relational Invariants, Not to Environmental or Evolutionary Novelty. The Journal of Mind and Behavior, 38 (2), 119-153.
Matzel, L. D., & Kolata, S. (2010). Selective attention, working memory, and animal intelligence. Neuroscience & Biobehavioral Reviews, 34 (1), 23-30.
O'keefe, J., & Nadel, L. (1978). The hippocampus as a cognitive map. Oxford university press.
Van Hoesen, G. W., & Pandya, D. N. (1975). Some connections of the entorhinal (area 28) and perirhinal (area 35) cortices of the rhesus monkey. III. Efferent connections. Brain research, 95(1), 39-59.
Waltz, J. A., Knowlton, B. J., Holyoak, K. J., Boone, K. B., Mishkin, F. S., de Menezes Santos, M., ... & Miller, B. L. (1999). A system for relational reasoning in human prefrontal cortex. Psychological science, 10(2), 119-125.
Zola-Morgan, S., Squire, L. R., Amaral, D. G., & Suzuki, W. A. (1989). Lesions of perirhinal and parahippocampal cortex that spare the amygdala and hippocampal formation produce severe memory impairment. Journal of Neuroscience, 9(12), 4355-4370.
Attributions
Section 10.7 adapted by Kenneth A. Koenigshofer, PhD., from The Brain from Top to Bottom; license: Copyleft, https://thebrain.mcgill.ca/flash/pop.../pop_copy.html. licensed under CC BY 4.0.