
11.3: Stages of Sleep
- Page ID
- 110610
This page is a draft and under active development. Please forward any questions, comments, and/or feedback to the ASCCC OERI (oeri@asccc.org).
- Explain the different stages of sleep.
- Describe the nature of REM sleep, its biological underpinnings and possible functions.
Overview
While watching someone sleep might appear as if they are seemingly unconscious with eyes closed for an extended period of time, in fact brain wave recordings show that the brain is cycling through several different stages as an individual sleeps. These stages each have distinct characteristics of brain activity and conscious experience and are divided into two main types known as Rapid Eye Movement (REM) and Non-REM. This section will explore these characteristics and some of the peculiar and unexpected features of REM sleep, in particular.
The different stages of sleep
From a behavioral standpoint, sleep is defined by four criteria:
- reduced motor activity
- diminished responses to external stimuli
- stereotyped posture (in humans, lying down with eyes closed)
- relatively ready reversibility.
These criteria distinguish sleep from hibernation and unconscious states, such as coma.
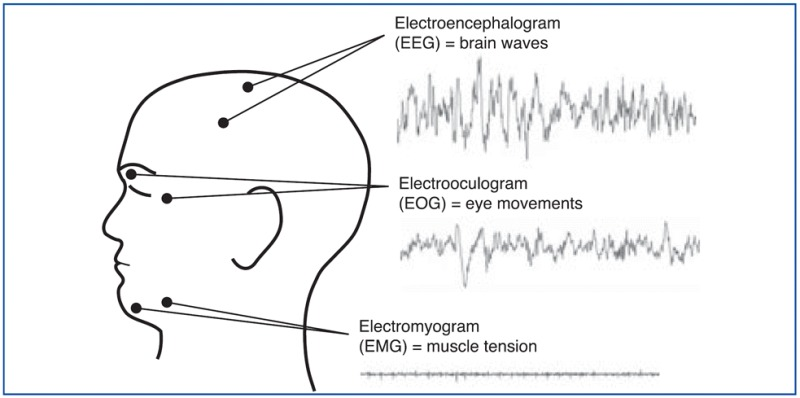
Sleep is a highly organized sequence of events that follows a regular, cyclic program each night. Polysomnograms - literally "many sleep measures" - are used to measure different body functions in a sleep lab (See Figure to see where electrodes are placed for each of the measurements). Thus, the EEG (electroencephalogram) is the technique used to measure sleep most readily and often (as discussed in chapter 2). For the EEG, electrodes are placed on the head and the brain's electrical activity is amplified and recorded). EMG (electromyograph that measures muscle tension), and EOG (electrooculogram that measures eye movement) patterns also change in predictable ways several times during a single sleep period.
NREM sleep is divided into four stages according to the amplitude and frequency of brain wave activity. In general, the EEG pattern of NREM sleep is slower, often more regular, and usually of higher voltage than that of wakefulness. As sleep gets deeper, the brain waves get slower and have greater amplitude. NREM Stage 1 is very light sleep; NREM Stage 2 has special brain waves called sleep spindles and K complexes; NREM Stages 3 and 4 show increasingly more high-voltage slow waves. In NREM Stage 4, it is extremely hard to be awakened by external stimuli. The muscle activity of NREM sleep is low, but the muscles retain their ability to function. Eye movements normally do not occur during NREM sleep, except for very slow eye movements, usually at the beginning. With the exception of brain activity, the body's general physiology during these stages is fairly similar to the wake state.
From the time you fall asleep to the time you reach the deepest non-REM sleep, about 1½ hours later, the amplitude of these waves increases continuously, while their frequency diminishes correspondingly. Compared with wakefulness and with REM (rapid eye movement) sleep, non-REM sleep is characterized by an electroencephalogram (EEG) in which the waves have a greater amplitude and a lower frequency.
Hypnograms were developed to summarize the voluminous chart recordings of electrical activities (EEG, EOG, and EMG) collected during a night's sleep. Hypnograms provide a simple way to display information originally collected on many feet of chart paper or stored as a large file on a computer. The figure illustrates a hypnogram. (Of course, it is important to keep in mind that individuals' hypnograms might be much less uniform than this relative "ideal" presented here.)
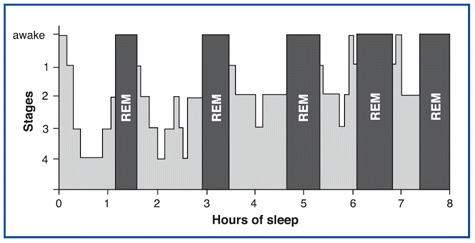
We can make several observations about the hypnogram in this figure. First, the periods of NREM and REM sleep alternate during the night. Second, the deepest stages of NREM sleep occur in the first part of the night. Third, the episodes of REM sleep are longer as the night progresses. This hypnogram also indicates two periods during the night when the individual awakened (at about six and seven hours into the night).
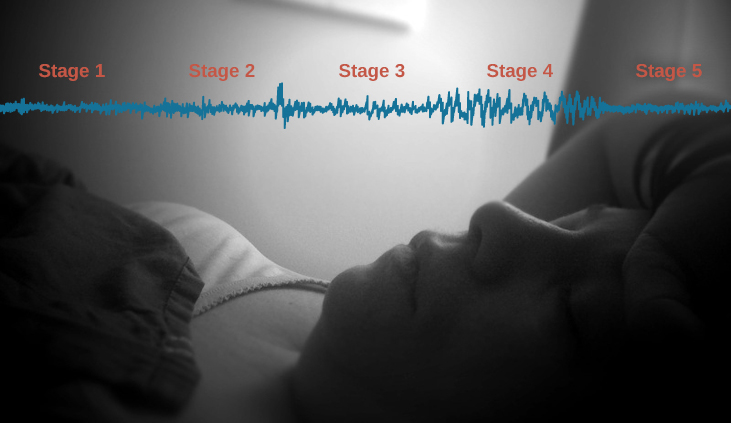
The four types of brain waves (shown in Figure \(\PageIndex{3}\), and others discussed below, are important criteria that have been used to define four distinct stages of non-REM (NREM) sleep. Obviously, falling into a deeper and deeper sleep as the night progresses is actually a gradual, continuous process, but these four stages still provide a convenient means of describing the relative depth of NREM sleep.
| Stage | EEG | Description |
|---|---|---|
| Stage 1 | 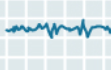 |
Stage 1 non-REM sleep begins when you first lie down and close your eyes. After a few sudden, sharp muscle contractions in the legs, the muscles relax. Then, as you continue falling asleep, the rapid beta waves of wakefulness are replaced by the slower alpha waves of someone who is relaxed with their eyes closed. Soon, the even slower theta waves begin to emerge. Though your reactions to stimuli from the outside world diminish, Stage 1 is still the phase of sleep from which it is easiest to wake someone up. In experiments where people are awakened from Stage 1 sleep and asked about their state of consciousness, they usually report that they had just fallen asleep or had been in the process of doing so. They also often report having had stray thoughts and short dreams. Each period of Stage 1 sleep generally lasts 3 to 12 minutes, |
| Stage 2 | 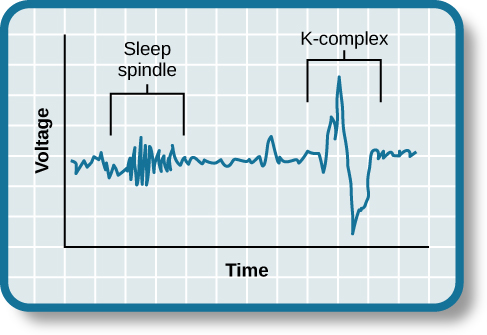 |
Stage 2 non-REM sleep is a stage of light sleep in which the frequency of the EEG trace decreases further while its amplitude increases. The theta waves characteristic of Stage 2 sleep are interrupted by occasional series of high-frequency waves known as sleep spindles. These bursts of activity have a frequency of 8 to 14 Hz and an amplitude of 50 to 150 µV. Sleep spindles generally last 1 to 2 seconds. They are generated by interactions between thalamic and cortical neurons. During Stage 2 sleep, the EEG trace may also show a fast, high-amplitude wave form called a K-complex. The K-complex seems to be associated with brief awakenings, often in response to external stimuli. People in Stage 2 sleep are unlikely to react to a light or a noise, unless it is extremely bright or loud. It is still possible to awaken them, even if they then report that they were really sleeping during the 10 to 20 minutes that this stage lasts during the earliest of the night’s sleep cycles. But because people go through Stage 2 sleep several times during the cycles in a night, this is the stage in which adults spend the greatest proportion of their sleep–nearly 50% of the total time that they sleep each night. |
| Stage 3 | 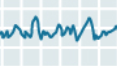 |
Stage 3 non-REM sleep marks the passage from moderately to truly deep sleep. Delta waves appear and soon account for nearly half of the waves in the EEG trace. Sleep spindles and K-complexes still occur, but less often than in Stage 2. The greater activity observed in the electro-oculogram (EOG) trace during stages 3 and 4 reflects the greater amplitude of EEG activity in the prefrontal areas, rather than movements of the eyes. Stage 3 lasts about 10 minutes during the first sleep cycle of the night but accounts for only about 7% of a total night’s sleep. During Stage 3, the muscles still have some tonus, and sleepers show very little response to external stimuli unless they are very strong or have a special personal meaning (for example, when someone calls your name, or when a baby cries within earshot of its mother). |
| Stage 4 | 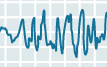 |
Stage 4 non-REM sleep is the deepest, the one in which we sleep the most soundly. The EEG trace is dominated by delta waves, and overall neuronal activity is at its lowest. The brain’s temperature is also at its lowest, and breathing, heart rate, and blood pressure are all reduced under the influence of the parasympathetic nervous system. In adults, Stage 4 lasts about 35 to 40 minutes during the first sleep cycle of the night; it accounts for 15 to 20% of total sleep time in young adults. The muscles still have their tonus, and some movements of the arms, legs, and trunk are possible. This is the stage of sleep that accomplishes most of the body’s repair work and from which it is most difficult to wake someone up. This is also the stage of sleep in which children may have episodes of somnambulism (sleepwalking) and night terrors. |
REM/NREM Sleep Chemicals and Brainstem Structures
In addition to maintaining wakefulness, several of the nuclei of the two pathways from the brainstem and the pons to the cortex use acetylcholine and glutamate as neurotransmitters and are partly responsible for the cortical activation that occurs during REM sleep.
The midbrain reticular formation projects massively into the thalamic nuclei, which in turn influence the entire cortex. The role of this formation is to desynchronize the cortex in the broad sense, thus facilitating not only wakefulness but REM sleep as well. Formerly known as the ascending activating reticular system, it is now regarded simply as part of the wakefulness network.
Brain structures involved in REM
Rapid eye movement (REM) sleep is a distinct, homeostatically controlled brain state characterized by an activated electroencephalogram (EEG) in combination with paralysis of skeletal muscles and is associated with vivid dreaming.
In 1953, Aserinsky and Kleitman first reported the existence of rapid eye movement (REM) sleep in humans as a periodically recurring brain state marked by a low amplitude electroencephalogram (EEG) and rapid eye movements (Aserinsky and Kleitman, 1953). Soon afterward, Kleitman and Dement found that the EEG during REM sleep resembles that during alert waking and showed that REM sleep coincides with periods of vivid dreaming (Dement and Kleitman, 1957). Two years later, Jouvet discovered in cats that the activated EEG during REM sleep is associated with a complete paralysis of skeletal muscles, reflected in a flat electromyogram (EMG), and therefore coined the term paradoxical sleep (Jouvet and Michel, 1959). Besides these defining properties in EEG and EMG, REM sleep is characterized by further striking neurophysiological and behavioral features, including high-amplitude theta oscillations in the hippocampus, muscle twitches, autonomic and respiratory activation, an elevated arousal threshold and bursts of large waves in the local field potential (LFP), called PGO-waves (P= pons, G= (lateral) geniculate nucleus, O= occipital cortex ), that originate in the pons and propagate to the lateral geniculate nucleus, occipital cortex, and other brain areas (Datta, 1997; Karashima et al., 2010).
The fact that the prefrontal cortex is relatively silent in REM falls in line with the fact that our dream content is often bizarre, illogical and socially inappropriate. Also the anterior cingulate gyrus which plays a role in attention and motivation is quite involved in REM sleep. And it appears that certain nuclei in the pons help trigger REM.
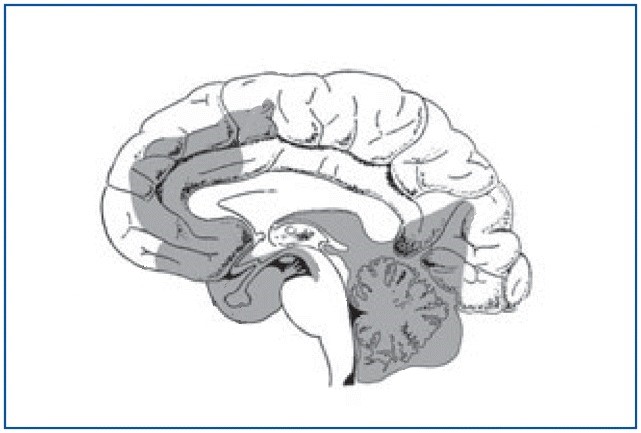
The amygdala is one of the parts of the brain that is most active during REM sleep, but this state is actually generated deep in the brainstem. However, the set of cortical and limbic structures involved in REM sleep do not just passively submit to orders issued by the brainstem. On the contrary, the particular kind of dream images associated with REM sleep are the result of a dynamic interaction between certain key structures in the brainstem and the rest of the brain (See Figure \(\PageIndex{4}\)).
REM and dreams
It was shown by 1950s that the cortex is as active in REM as when someone is awake – which is one reason why REM is also called paradoxical sleep.
In humans, sleep is punctuated by REM (rapid eye movement) sleep about every 90 min (Ohayon et al., 2004). This is when most dreaming occurs (Hobson, 2009). Although some forms of dreaming can occur during non-REM sleep, such dreams are quite different from REM dreams; non-REM dreams usually are related to plans or thoughts, and they lack the visual vividness and hallucinatory and delusory components of REM dreams (Roffwarg et al., 1962; Nielsen, 2000). Rapid eye movement during sleep is thought to be associated with visual experience of dreaming (Andrillon et al., 2015).
It was also found using MRIs that while the primary visual cortex was relatively silent in REM (clearly their eyes are closed and therefore they aren’t seeing anything), the secondary visual areas of the cortex, that interpret and analyze visual information, are relatively active in REM state. This is consistent with the fact that when we are awakened from REM sleep we often report highly elaborate visual dream scenes.
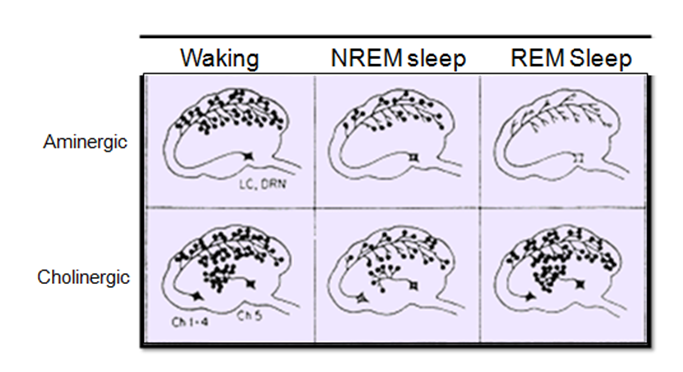
Neurochemical bases of REM
REM is a combination of increase in cholinergic (acetylcholine) activity (in pontine tegmentum/pons and midbrain) and decrease in aminergic (monoamines - serotonin and norepinephrine) activity (in dorsal raphe nucleus and locus coeruleus).
That a sudden elevation in the activity of the cholinergic neurons of the pons is necessary for the onset of REM sleep may seem somewhat strange, because the brain’s acetylcholine systems are known for their role in wakefulness. How then can this system associated with wakefulness be activated during one of the phases of sleep? The answer probably lies in the simultaneous reduction in the activity of two other nuclei that produce other wakefulness neurotransmitters, a reduction that is just as necessary for REM sleep as the increase in cholinergic activity. The two nuclei in question are the dorsal raphe nucleus, a group of serotonergic neurons, and the locus coeruleus, a group of noradrenergic neurons, and both of them are located in the rostral portion of the pons.
REM sleep cannot begin unless all activity in these two main aminergic systems of the brainstem ceases (See Figure \(\PageIndex{5}\) for some of the comparisons of neurochemical activation) . The serotonergic and noradrenergic neurons in these systems are referred to as REM-off neurons because of their inactivity during REM sleep, and they can be said to act as a sort of permissive system for REM sleep, in the sense that REM sleep becomes possible only when their activity has ceased. The shutdown of these neurons might, for example, help to suppress consciousness during REM sleep. Also, the periods of REM sleep end when these aminergic neurons become active again.
The rapid eye movements - that REM gets named for - come from originating signals in the pontine reticular formation sending signals to the midbrain and which coordinate the duration and direction of eye movements.
Musculature in REM
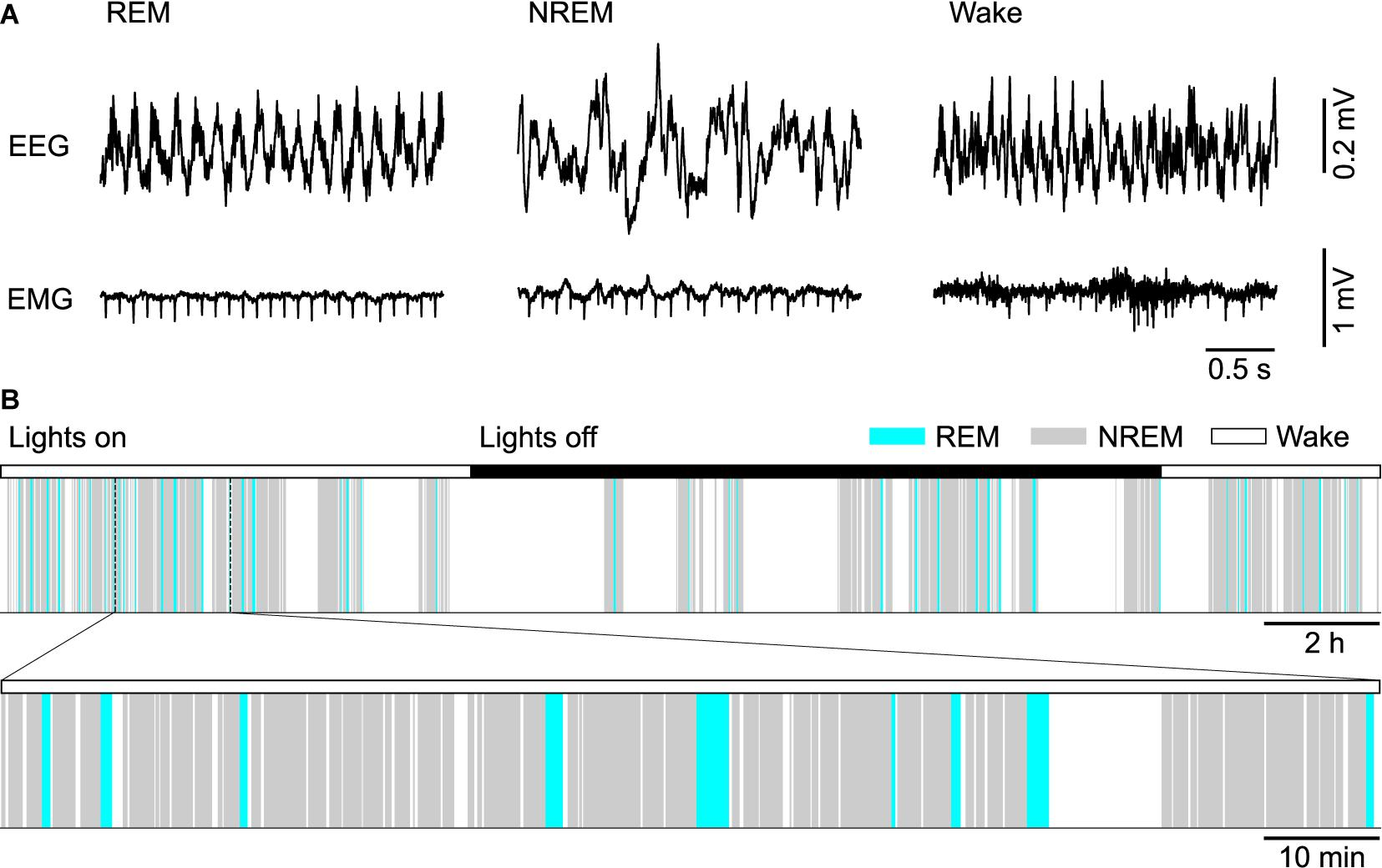
One of the most singular characteristics of REM sleep—the paralysis of the large muscles of the body that it causes —also is explained by phenomena that occur in certain parts of the brainstem. Intense neural activity of REM sleep excites majority of neurons in the cortex including those in the primary motor cortex. These generate organized sequences of activities that represent commands for bodily movements. But they never reach motor neurons of arms and legs (only the respiratory muscles, those of eye and middle ear actually respond to those commands). See Figure \(\PageIndex{6}\) for the coordination of EEG and EMG in the 24-hour hypnogram of a mouse. Increase of cholinergic activity in the pons will, ultimately, inhibit motor neurons in the spinal cord.
When you are awake, your brain’s wakefulness circuits exert controls that prevent you from displaying the forms of brain activity that characterize REM sleep. But in the human fetus, these controls are not yet in place, which may explain why, during the last few months of gestation, the fetus spends such a large proportion of its sleeping time (about 80%) in REM sleep.
PGO Waves - (Pons - geniculate - occipital cortex)
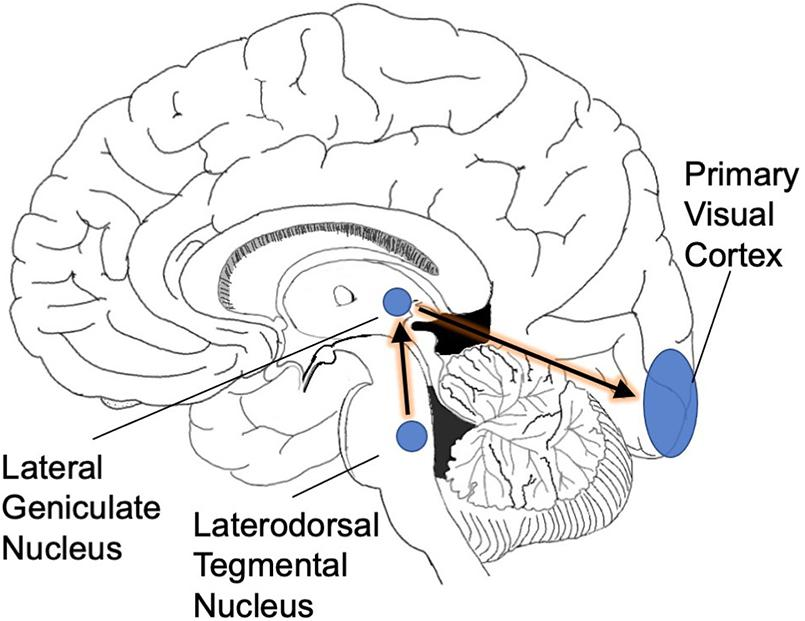
PGO waves are among the various phasic events that occur during REM sleep, along with the rapid eye movements and changes in breathing and heart rates. PGO waves can be generated in the absence of REM sleep by stimulation of the pons with acetylcholine. As shown in figure\(\PageIndex{7}\), the pons send signals to the thalamus which in turn signal the occipital cortex.
REM sleep is triggered by a specialized set of neurons in the pons (Hobson et al., 1975). Increased activity in this neuronal population has two consequences. First, elaborate neural circuitry keeps the body immobile during REM sleep by paralyzing major muscle groups (Chase, 2008). The muscle shut-down allows the brain to simulate a visual experience without moving the body at the same time. Second, we experience vision when waves of activity travel from the pons to the lateral geniculate nucleus and then to the occipital cortex (these are known as ponto-geniculo-occipital waves or PGO waves) (Gott et al., 2017). When the spikes of activity arrive at the occipital pole, we feel as though we are seeing even though our eyes are closed (Nir and Tononi, 2010). The visual cortical activity is presumably why dreams are pictorial and filmic instead of conceptual or abstract.
Summary
In an 8 hour typical night's sleep we move through distinct stages of sleep, several times through the night. We go from relative wakefulness to the deepest stages of sleep, cycle back out and into REM multiple times. Each of these stages correspond with different EEG wave patterns as well as the involvement of different regions of the brain and neurochemicals. The nature of REM sleep is paradoxical in a number of ways, and it seems to be different from the other stages in terms of quality and quantity.
References
- Aserinsky, E., and Kleitman, N. (1953). Regularly occurring periods of eye motility, and concomitant phenomena, during sleep. Science 118, 273–274. doi: 10.1126/science.118.3062.273
- Chase, M. H. (2008). Confirmation of the consensus that glycinergic postsynaptic inhibition is responsible for the atonia of REM sleep. Sleep 31, 1487–1491. doi: 10.1093/sleep/31.11.1487
- Datta, S. (1997). Cellular basis of pontine ponto-geniculo-occipital wave generation and modulation. Cell. Mol. Neurobiol. 17, 341–365.
- Dement, W., and Kleitman, N. (1957). The relation of eye movements during sleep to dream activity: an objective method for the study of dreaming. J. Exp. Psychol. 53, 339–346. doi: 10.1037/h0048189
- Gott, J. A., Liley, D. T. J., and Hobson, J. A. (2017). Towards a functional understanding of PGO Waves. Front. Hum. Neurosci. 11:89.
- Hobson, J. A., McCarley, R. W., and Wyzinski, P. W. (1975). Sleep cycle oscillation: reciprocal discharge by two brainstem neuronal groups. Science 189, 55–58. doi: 10.1126/science.1094539
- Jouvet, M., and Michel, F. (1959). Electromyographic correlations of sleep in the chronic decorticate & mesencephalic cat. C. R. Séances Soc. Biol. Ses Fil. 153, 422–425.
- Karashima, A., Katayama, N., and Nakao, M. (2010). Enhancement of synchronization between hippocampal and amygdala theta waves associated with pontine wave density. J. Neurophysiol. 103, 2318–2325. doi: 10.1152/jn.00551.2009
- Nir, Y., and Tononi, G. (2010). Dreaming and the brain: from phenomenology to neurophysiology. Trends Cogn. Sci. 14, 88–100. doi: 10.1016/j.tics.2009.12.001
Attributions
- Different types of sleep - intermediate by Bruno Dubuc under a Copyleft license.
- Different types of sleep - advanced by Bruno Dubuc under a Copyleft license.
- Adapted from Information about sleep by NIH in the public domain.
- Information about sleep by NIH in the public domain.
- The Brain structures that keep you up and put you to sleep - intermediate by Bruno Dubuc under a Copyleft license.
- The Brain structures that keep you up and put you to sleep - advanced by Bruno Dubuc under a Copyleft license.
- Neural and Homeostatic Regulation of REM sleep by Park and Weber, in Frontiers licensed CC-BY
- Stages of Sleep by Lumen Learning, authored by OpenStax, licensed CC BY 4.0
- The Defensive Activation Theory: REM Sleep as a Mechanism to Prevent the Takeover of the Visual Cortex by David Eagleman and Don Vaughn licensed CC-BY 4.0
- REM sleep (paradoxical sleep) characteristics by Dr. Mohammad Torabi Nami and Dr. Khsoro Sadeghniaat. Licensed CC BY 3.0 via Webmed Central