
11.2: Race, A Discredited Concept
- Last updated
- Save as PDF
- Page ID
- 56472
At some point in your life, you have probably been asked to identify your race on a college form, job application, government or military form, or some other official document. And most likely, you were required to select from a list of choices rather than given the ability to respond freely. The frequency with which we are exposed to four or five common racial labels—“white,” “black,” “Caucasian,” and “Asian,” for example—tends to promote the illusion that racial categories are natural, objective, and evident divisions. After all, if Justin Timberlake, Jay-Z, and Jackie Chan stood side by side, those common racial labels might seem to make sense. What could be more objective, more conclusive, than this evidence before our very eyes? By this point, you might be thinking that anthropologists have gone completely insane in denying biological human races!
Physical anthropologists have identified several important concepts regarding the true nature of humans’ physical, genetic, and biological variation that have discredited race as a biological concept. Many of the issues presented in this section are discussed in further detail in Race: Are We So Different, a website created by the American Anthropological Association. The American Anthropological Association (AAA) launched the website to educate the public about the true nature of human biological and cultural variation and challenge common misperceptions about race. This is an important endeavor because race is a complicated, often emotionally charged topic, leading many people to rely on their personal opinions and hearsay when drawing conclusions about people who are different from them. The website is highly interactive, featuring multimedia illustrations and online quizzes designed to increase visitors’ knowledge of human variation. I encourage you to explore the website as you will likely find answers to several of the questions you may still be asking after reading this chapter.[3]
Before explaining why distinct biological races do not exist among humans, I must point out that one of the biggest reasons so many people continue to believe in the existence of biological human races is that the idea has been intensively reified in literature, the media, and culture for more than three hundred years. Reification refers to the process in which an inaccurate concept or idea is so heavily promoted and circulated among people that it begins to take on a life of its own. Over centuries, the notion of biological human races became ingrained—unquestioned, accepted, and regarded as a concrete “truth.” Studies of human physical and cultural variation from a scientific and anthropological perspective have allowed us to move beyond reified thinking and toward an improved understanding of the true complexity of human diversity.
Definition: reified
The process by which an inaccurate concept or idea is accepted as “truth.”
The reification of race has a long history. Especially during the eighteenth and nineteenth centuries, philosophers and scholars attempted to identify various human races. They perceived “races” as specific divisions of humans who shared certain physical and biological features that distinguished them from other groups of humans. This historic notion of race may seem clear-cut and innocent enough, but it quickly led to problems as social theorists attempted to classify people by race. One of the most basic difficulties was the actual number of human races: how many were there, who were they, and what grounds distinguished them? Despite more than three centuries of such effort, no clear-cut scientific consensus was established for a precise number of human races.

One of the earliest and most influential attempts at producing a racial classification system came from Swedish botanist Carolus Linnaeus, who argued in Systema Naturae (1735) for the existence of four human races: Americanus (Native American / American Indian), Europaeus (European), Asiaticus (East Asian), and Africanus (African). These categories correspond with common racial labels used in the United States for census and demographic purposes today. However, in 1795, German physician and anthropologist Johann Blumenbach suggested that there were five races, which he labeled as Caucasian (white), Mongolian (yellow or East Asian), Ethiopian (black or African), American (red or American Indian), Malayan (brown or Pacific Islander). Importantly, Blumenbach listed the races in this exact order, which he believed reflected their natural historical descent from the “primeval” Caucasian original to “extreme varieties.”[4] Although he was a committed abolitionist, Blumenbach nevertheless felt that his “Caucasian” race (named after the Caucasus Mountains of Central Asia, where he believed humans had originated) represented the original variety of humankind from which the other races had degenerated.
By the early twentieth century, many social philosophers and scholars had accepted the idea of three human races: the so-called Caucasoid, Negroid, and Mongoloid groups that corresponded with regions of Europe, sub-Saharan Africa, and East Asia, respectively. However, the three-race theory faced serious criticism given that numerous peoples from several geographic regions were omitted from the classification, including Australian Aborigines, Asian Indians, American Indians, and inhabitants of the South Pacific Islands. Those groups could not be easily pigeonholed into racial categories regardless of how loosely the categories were defined. Australian Aborigines, for example, often have dark complexions (a trait they appeared to share with Africans) but reddish or blondish hair (a trait shared with northern Europeans). Likewise, many Indians living on the Asian subcontinent have complexions that are as dark or darker than those of many Africans and African Americans. Because of these seeming contradictions, some academics began to argue in favor of larger numbers of human races—five, nine, twenty, sixty, and more.[5]
During the 1920s and 1930s, some scholars asserted that Europeans were comprised of more than one “white” or “Caucasian” race: Nordic, Alpine, and Mediterranean (named for the geographic regions of Europe from which they descended). These European races, they alleged, exhibited obvious physical traits that distinguished them from one another and thus served as racial boundaries. For example, “Nordics” were said to consist of peoples of Northern Europe—Scandinavia, the British Isles, and Northern Germany— while “Alpines” came from the Alps Mountains of Central Europe and included French, Swiss, Northern Italians, and Southern Germans. People from southern Europe—including Portuguese, Spanish, Southern Italians, Sicilians, Greeks, and Albanians—comprised the “Mediterranean” race. Most Americans today would find this racial classification system bizarre, but its proponents argued for it on the basis that one would observe striking physical differences between a Swede or Norwegian and a Sicilian. Similar efforts were made to “carve up” the populations of Africa and Asia into geographically local, specific races.[6]
The fundamental point here is that any effort to classify human populations into racial categories is inherently arbitrary and subjective rather than scientific and objective. These racial classification schemes simply reflected their proponents’ desires to “slice the pie” of human physical variation according to the particular trait(s) they preferred to establish as the major, defining criteria of their classification system. Two major types of “race classifiers” have emerged over the past 300 years: lumpers and splitters. Lumpers have classified races by large geographic tracts (often continents) and produced a small number of broad, general racial categories, as reflected in Linnaeus’s original classification scheme and later three-race theories. Splitters have subdivided continent-wide racial categories into specific, more localized regional races and attempted to devise more “precise” racial labels for these specific groups, such as the three European races described earlier. Consequently, splitters have tried to identify many more human races than lumpers.
Racial labels, whether from a lumper or a splitter model, clearly attempt to identify and describe something. So why do these racial labels not accurately describe human physical and biological variation? To understand why, we must keep in mind that racial labels are distinct, discrete categories while human physical and biological variations (such as skin color, hair color and texture, eye color, height, nose shape, and distribution of blood types) are continuous rather than discrete.
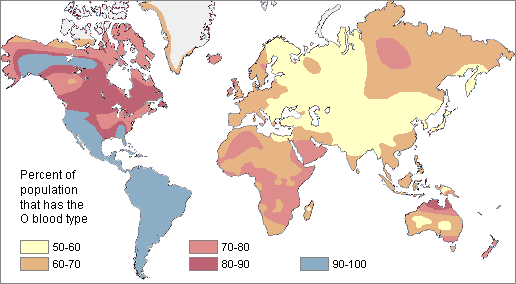
Physical anthropologists use the term cline to refer to differences in the traits that occur in populations across a geographical area. In a cline, a trait may be more common in one geographical area than another, but the variation is gradual and continuous with no sharp breaks. A prominent example of clinal variation among humans is skin color. Think of it this way: Do all “white” persons who you know actually share the same skin complexion? Likewise, do all “black” persons who you know share an identical skin complexion? The answer, obviously, is no, since human skin color does not occur in just 3, 5, or even 50 shades. The reality is that human skin color, as a continuous trait, exists as a spectrum from very light to very dark with every possible hue, shade, and tone in between.
Definition: cline
Differences in the traits that occur in populations across a geographical area. In a cline, a trait may be more common in one geographical area than another, but the variation is gradual and continuous, with no sharp breaks.
Imagine two people—one from Sweden and one from Nigeria—standing side by side. If we looked only at those two individuals and ignored people who inhabit the regions between Sweden and Nigeria, it would be easy to reach the faulty conclusion that they represented two distinct human racial groups, one light (“white”) and one dark (“black”). [7] However, if we walked from Nigeria to Sweden, we would gain a fuller understanding of human skin color because we would see that skin color generally became gradually lighter the further north we traveled from the equator. At no point during this imaginary walk would we reach a point at which the people abruptly changed skin color. As physical anthropologists such as John Relethford (2004) and C. Loring Brace (2005) have noted, the average range of skin color gradually changes over geographic space. North Africans are generally lighter-skinned than Central Africans, and southern Europeans are generally lighter-skinned than North Africans. In turn, northern Italians are generally lighter-skinned than Sicilians, and the Irish, Danes, and Swedes are generally lighter-skinned than northern Italians and Hungarians. Thus, human skin color cannot be used as a definitive marker of racial boundaries.
There are a few notable exceptions to this general rule of lighter-complexioned people inhabiting northern latitudes. The Chukchi of Eastern Siberia and Inuits of Alaska, Canada, and Greenland have darker skin than other Eurasian people living at similar latitudes, such as Scandinavians. Physical anthropologists have explained this exception in terms of the distinct dietary customs of indigenous Arctic groups, which have traditionally been based on certain native meats and fish that are rich in Vitamin D (polar bears, whales, seals, and trout).
What does Vitamin D have to do with skin color? The answer is intriguing! Dark skin blocks most of the sun’s dangerous ultraviolet rays, which is advantageous in tropical environments where sunlight is most intense. Exposure to high levels of ultraviolet radiation can damage skin cells, causing cancer, and also destroy the body’s supply of folate, a nutrient essential for reproduction. Folate deficiency in women can cause severe birth defects in their babies. Melanin, the pigment produced in skin cells, acts as a natural sunblock, protecting skin cells from damage, and preventing the breakdown of folate. However, exposure to sunlight has an important positive health effect: stimulating the production of vitamin D. Vitamin D is essential for the health of bones and the immune system. In areas where ultraviolet radiation is strong, there is no problem producing enough Vitamin D, even as darker skin filters ultraviolet radiation.[8]
In environments where the sun’s rays are much less intense, a different problem occurs: not enough sunlight penetrates the skin to enable the production of Vitamin D. Over the course of human evolution, natural selection favored the evolution of lighter skin as humans migrated and settled farther from the equator to ensure that weaker rays of sunlight could adequately penetrate our skin. The diet of indigenous populations of the Arctic region provided sufficient amounts of Vitamin D to ensure their health. This reduced the selective pressure toward the evolution of lighter skin among the Inuit and the Chukchi. Physical anthropologist Nina Jablonski (2012) has also noted that natural selection could have favored darker skin in Arctic regions because high levels of ultraviolet radiation from the sun are reflected from snow and ice during the summer months.
Still, many people in the United States remain convinced that biologically distinct human races exist and are easy to identify, declaring that they can walk down any street in the United States and easily determine who is “white” and who is “black.” The United States was populated historically by immigrants from a small number of world regions who did not reflect the full spectrum of human physical variation. The earliest settlers in the North American colonies overwhelmingly came from Northern Europe (particularly, Britain, France, Germany, and Ireland), regions where skin colors tend to be among the lightest in the world. Slaves brought to the United States during the colonial period came largely from the western coast of Central Africa, a region where skin color tends to be among the darkest in the world. Consequently, when we look at today’s descendants of these groups, we are not looking at accurate, proportional representations of the total range of human skin color; instead, we are looking, in effect, at opposite ends of a spectrum, where striking differences are inevitable. More recent waves of immigrants who have come to the United States from other world regions have brought a wider range of skin colors, shaping a continuum of skin color that defies classification into a few simple categories.
Physical anthropologists have also found that there are no specific genetic traits that are exclusive to a “racial” group. For the concept of human races to have biological significance, an analysis of multiple genetic traits would have to consistently produce the same racial classifications. In other words, a racial classification scheme for skin color would also have to reflect classifications by blood type, hair texture, eye shape, lactose intolerance, and other traits often mistakenly assumed to be “racial” characteristics. An analysis based on any one of those characteristics individually would produce a unique set of racial categories because variations in human physical and genetic are nonconcordant. Each trait is inherited independently, not “bundled together” with other traits and inherited as a package. There is no correlation between skin color and other characteristics such as blood type and lactose intolerance.
Definition: nonconcordant
Genetic traits that are inherited independently rather than as a package.
A prominent example of nonconcordance is sickle-cell anemia, which people often mistakenly think of as a disease that only affects Africans, African Americans, and “black” persons. In fact, the sickle-cell allele (the version of the gene that causes sickle-cell anemia when a person inherits two copies) is relatively common among people whose ancestors are from regions where a certain strain of malaria, plasmodium falciparum, is prevalent, namely Central and Western Africa and parts of Mediterranean Europe, the Arabian peninsula, and India. The sickle-cell trait thus is not exclusively African or “black.” The erroneous perceptions are relatedly primarily to the fact that the ancestors of U.S. African Americans came predominantly from Western Africa, where the sickle-cell gene is prevalent, and are therefore more recognizable than populations of other ancestries and regions where the sickle-cell gene is common, such as southern Europe and Arabia.[9]
Another trait commonly mistaken as defining race is the epicanthic eye fold typically associated with people of East Asian ancestry. The epicanthic eye fold at the outer corner of the eyelid produces the eye shape that people in the United States typically associate with people from China and Japan, but is also common in people from Central Asia, parts of Eastern Europe and Scandinavia, some American Indian groups, and the Khoi San of southern Africa.
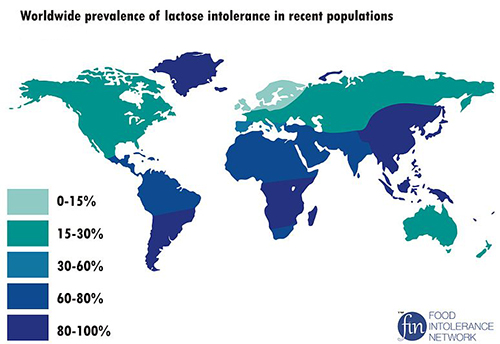
In college, I took a course titled “Nutrition” because I thought it would be an easy way to boost my grade point average. The professor of the class, an authoritarian man in his late 60s or early 70s, routinely declared that “Asians can’t drink milk!” When this assertion was challenged by various students, including a woman who claimed that her best friend was Korean and drank milk and ate ice cream all the time, the professor only became more strident, doubling down on his dairy diatribe and defiantly vowing that he would not “ignore the facts” for “purposes of political correctness.” However, it is scientific accuracy, not political correctness, we should be concerned about, and lactose tolerance is a complex topic. Lactose is a sugar that is naturally present in milk and dairy products, and an enzyme, lactase, breaks it down into two simpler sugars that can be digested by the body. Ordinarily, humans (and other mammals) stop producing lactase after infancy, and approximately 75 percent of humans are thus lactose intolerant and cannot naturally digest milk. Lactose intolerance is a natural, normal condition. However, some people continue to produce lactase into adulthood and can naturally digest milk and dairy products. This lactose persistence developed through natural selection, primarily among people in regions that had long histories of dairy farming (including the Middle East, Northern Europe, Eastern Europe, East Africa, and Northern India). In other areas and for some groups of people, dairy products were introduced relatively recently (such as East Asia, Southern Europe, and Western and Southern Africa and among Australian Aborigines and American Indians) and lactose persistence has not developed yet.[10]
The idea of biological human races emphasizes differences, both real and perceived, between groups and ignores or overlooks differences within groups. The biological differences between “whites” and “blacks” and between “blacks” and “Asians” are assumed to be greater than the biological differences among “whites” and among “blacks.” The opposite is actually true; the overwhelming majority of genetic diversity in humans (88–92 percent) is found within people who live on the same continent.[11] Also, keep in mind that human beings are one of the most genetically similar of all species. There is nearly six times more genetic variation among white-tailed deer in the southern United States than in all humans! Consider our closest living relative, the chimpanzee. Chimpanzees’ natural habitat is confined to central Africa and parts of western Africa, yet four genetically distinct groups occupy those regions and they are far more genetically distinct than humans who live on different continents. That humans exhibit such a low level of genetic variation compared to other species reflects the fact that we are a relatively recent species; modern humans (Homo sapiens) first appeared in East Africa just under 200,000 years ago.[12]
Physical anthropologists today analyze human biological variation by examining specific genetic traits to understand how those traits originated and evolved over time and why some genetic traits are more common in certain populations. Since much of our biological diversity occurs mostly within (rather than between) continental regions once believed to be the homelands of distinct races, the concept of race is meaningless in any study of human biology. Franz Boas, considered the father of modern U.S. anthropology, was the first prominent anthropologist to challenge racial thinking directly during the early twentieth century. A professor of anthropology at Columbia University in New York City and a Jewish immigrant from Germany, Boas established anthropology in the United States as a four-field academic discipline consisting of archaeology, physical/biological anthropology, cultural anthropology, and linguistics. His approach challenged conventional thinking at the time that humans could be separated into biological races endowed with unique intellectual, moral, and physical abilities.
In one of his most famous studies, Boas challenged craniometrics, in which the size and shape of skulls of various groups were measured as a way of assigning relative intelligence and moral behavior. Boas noted that the size and shape of the skull were not fixed characteristics within groups and were instead influenced by the environment. Children born in the United States to parents of various immigrant groups, for example, had slightly different average skull shapes than children born and raised in the homelands of those immigrant groups. The differences reflected relative access to nutrition and other socio-economic dimensions. In his famous 1909 essay “Race Problems in America,” Boas challenged the commonly held idea that immigrants to the United States from Italy, Poland, Russia, Greece, the Austro-Hungarian Empire, and other southern and eastern European nations were a threat to America’s “racial purity.” He pointed out that the British, Germans, and Scandinavians (popularly believed at the time to be the “true white” heritages that gave the United States its superior qualities) were not themselves “racially pure.” Instead, many different tribal and cultural groups had intermixed over the centuries. In fact, Boas asserted, the notion of “racial purity” was utter nonsense. As present-day anthropologist Jonathan Marks (1994) noted, “You may group humans into a small number of races if you want to, but you are denied biology as a support for it.”[13]
BIBLIOGRAPHY
Boas, Franz. “Race Problems in America.” Science 29 no. 752 (1909): 839–849.
Brace, C. Loring. ‘Race’ is a Four-Letter Word: The Genesis of the Concept. New York: Oxford University Press, 2005.
Jablonski, Nina. Living Color: The Biological and Social Meaning of Skin Color. Berkeley, CA: University of California Press, 2012.
Marks, Jonathan. “Black, White, Other.” Natural History December, 1994: 32–35.
Relethford, John H. Reflections Of Our Past: How Human History Is Revealed In Our Genes. Boulder, CO: Westview Press, 2004.
NOTES
- More discussion of the material in this section can be found in Carol Mukhopadhyay, Rosemary Henze, and Yolanda Moses, How Real Is Race? A Sourcebook on Race, Culture, and Biology (Lanham, MD: Rowman & Littlefield, 2013). Chapters 5 and 6 discuss the cultural construction of racial categories as a form of classification. The Race: Are We So Different website and its companion resources for teachers and researchers also explore the ideas described here. ↵
- Johann Friedrich Blumenbach, On the Natural Varieties of Mankind: De Generis Humani Varietate Nativa (New York: Bergman Publishers, 1775). ↵
- For details about how these categories were established, see Stephen Jay Gould, The Mismeasure of Man. ↵
- For a discussion of the efforts to subdivide racial groups in the nineteenth century and its connection to eugenics, see Carol Mukhopadhyay, Rosemary Henze, and Yolanda Moses, How Real Is Race? A Sourcebook on Race, Culture, and Biology.↵
- For more information about the genetic variation between human groups that puts this example in context see Sheldon Krimsky and Kathleen Sloan, Race and the Genetic Revolution: Science, Myth, and Culture (New York: Columbia University Press, 2011), 174-180. ↵
- Carol Mukhopadhyay et. al How Real Is Race? A Sourcebook on Race, Culture, and Biology, 43-48. ↵
- Ibid., 50-52. ↵
- Ibid., 50-51. ↵
- Ibid., 62. ↵
- Alan R. Templeton, “Human Races: A Genetic and Evolutionary Perspective” American Anthropologist 100 no. 3 (1998): 632-650. ↵
- Jonathan Marks, “Black, White, Other,” 35. ↵
Adapted From
"Race and Ethnicity" by Justin D. García, Millersville University of Pennsylvania. In Perspectives: An Open Invitation to Cultural Anthropology, 2nd Edition, Society for Anthropology in Community Colleges, 2020, under CC BY-NC 4.0.