
3.3: Homo Genus
- Last updated
- Save as PDF
- Page ID
- 4955

The emergence of the genus Homo marks the advent of larger brains, the emergence of material culture (at least material culture that survives in the archaeological record), and the eventual colonization of the world outside of Africa. The earliest Homo species are contemporaneous with several australopiths: Au. africanus, Au. garhi, Au. sediba, and all of the Paranthropus species. Africa was flush with hominins (Figure \(\PageIndex{0}\)). There are several trends we see in the evolution of the earliest Homo species to ourselves, Homo sapiens:
- Rounding of the cranium
- Enlargement and rewiring of the brain
- Smaller faces and teeth
- Decreasing prognathism
- Tallness
- Diversity of cultural traits
As with the overview on early hominins, we will continue taking the lumper approach. Several Homo species will not be discussed in detail, but may be mentioned in passing, e.g., Homo rudolfensis, Homo ergaster, Homo gautengensis, Homo antecessor, Homo cepranensis, Homo rhodesiensis, Homo tsaichangensis. We will first examine the morphological characteristics of various Homo species. Cultural traits will be addressed separately.
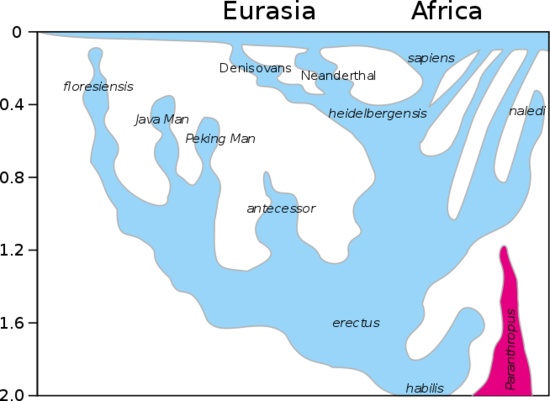
Figure \(\PageIndex{0}\): A model of the evolution of the genus Homo over the last 2 million years (vertical axis). The rapid "Out of Africa" expansion of H. sapiens is indicated at the top of the diagram, with admixture indicated with Neanderthals, Denisovans, and unspecified archaic African hominins. Late survival of robust australopithecines (Paranthropus) alongside Homo until 1.2 Mya is indicated in purple.
Homo habilis
Homo habilis was first discovered by Louis and Mary Leakey at Olduvai Gorge, Tanzania in 1960. Associated with stone tools (Oldowan), the Leakeys named their discover “handy man.” H. habilis fossils have been found in Tanzania, Kenya, Ethiopia, and South Africa, although there is some debate as to whether the South Africa specimens should be included in the species. Some paleoanthropologist contend that there was another early Homo species, Homo rudolfensis, which date back to 2.4mya. The H. rudolfensis fossils are slightly larger than those of H. habilis, leading some researchers to suggest the H. habilis exhibited sexual dimorphism and what we are seeing are male and female specimens of H. habilis. Others claim the size differences are significant enough to warrant the two species designations (O’Neil c1999-2012). In 2013 a Homo mandible was discovered in the Ledi-Geraru research area, Afar, Ethiopia. Dated to 2.8 to 2.75mya, the mandible exhibits an Australopithecus-like chin and Homo-like teeth (Villmoare et al. 2015). While still early in the research process, this discovery and further research may push back the date of the origin of Homo and help to resolve the debate between the H. rudolfensis and H. habilis fossils. For our purpose, we will consider them all H. habilis, making the date range for this hominin 2.4-1.4mya.
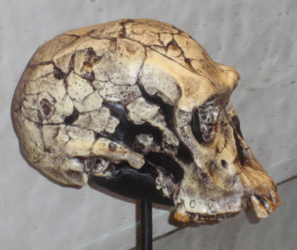
Figure \(\PageIndex{1}\) - Homo habilis
Morphologically, H. habilis has a larger brain than the australopiths, about 35% larger (O’Neil c1999-2012). You will recall from the section on trends in human evolution that the brain also began to rewire at this point. H. erectus exhibits less prognathism and platycephaly than early hominins. The brow ridge is also smaller. All of these traits together make the face smaller than the australopiths. Postcranially, H. habilis exhibit a mix of primitive and derived traits. Primitive traits connecting it to an australopith ancestor are the longer forearms and the size of the finger bones along with how the tendons attach to the wrist bones. The tips of the finger bones are broad like humans. Smaller teeth, a dental arcade in the shape of a parabolic arch, foot morphology, and a more rounded skull complete the human-like traits. Microanalysis of tooth wear indicates that H. habilis was omnivorous.
Homo erectus
Based on current fossil data, Homo erectus existed between 1.9mya to 25 kya (thousand years ago) (Jurmain 2013). Eugene Dubois found the first H. erectus, literally “upright human,” fossil in 1891 in Trinil, Java. H. erectus fossils have also been found in Africa, China, Europe, and Israel. Based on morphological differences in the cranium, some scientists identify two species, H. erectus in Asia and H. ergaster in Africa with the African specimens being smaller than the Asian; however, we will use the H. erectus designation.

Figure \(\PageIndex{2}\) - Homo erectus: Turkana Boy
H. erectus is the first hominin to reach modern human height. In 1984, a nearly complete H. erectus skeleton was found along the Nariokotome River in northern Kenya. Potassium-argon dating places “Turkana Boy” between 1.64 and 1.33mya. Aging and sexing of the fossil remains indicate that the individual was a male about age eight. He stood about 5’3” tall. If H. erectus followed the growth pattern of modern humans then Turkana Boy could have reached 6’ as an adult; however recent studies indicate that Turkana Boy followed the growth pattern of apes, so would have been near his adult height at the time of his death (Jurmain et al. 2013).
The pattern of increased brain size continued with H. erectus; its brain is up to 50% larger than its predecessor (O’Neil c1999-2012). This large brain was supported by a diet heavy in meat and other proteins. Its distinguishing characteristics include its sagittal keeling (a thickening of bone that runs from front to back on top of the skull) massive brow ridges (supraorbital tori) and bony prominences on the back of the skull. These prominences give the back of H. erectus’ skull a pentagon shape from behind. From the side, its skull is football-shaped. Postcranially, H. erectus’ bones are thicker than H. habilis, as were its jaws and face bones, and the proportion of arms to legs is lie that of modern humans, causing some to suggest that its bipedal gait was like ours. The length of its leg bones indicates that H. erectus would have been an efficient long-distance runner, allowing them to “run down small and even medium size game animals on the tropical savannas of East Africa” (O’Neil c1999-2012). If so, it is likely that H. erectus had much less body hair then its predecessors, as they would have needed to be able to sweat efficiently. Based on studies of human body lice, we know that head lice and pubic hair lice, or crab lice, are separate species. Crab lice are actually more closely related to gorilla hair lice then human head hair lice. The loss of body hair created a barrier between the two species, allowing them both to thrive on humans. DNA studies of the two species indicate that they split 3.0mya, suggesting that humans started to lose their body hair at that time. It is possible that H. erectus had little body hair (NOVA 2011).
Scientists generally agree that H. erectus was the first hominin to leave Africa. As mentioned previously, fossils have been found in Africa, China, Europe, and Israel. At Dmanisi, Republic of Georgia, fossils were found in strata dated to 1.7mya suggesting that H. erectus left Africa soon after it evolved. A recent report (Dembo et al. 2015) posits that H. habilis was the first hominin to leave Africa, not H. erectus. Should this contention be supported with more data, it can still be argued that H. erectus was quite successful in colonizing the Old World, helped, no doubt, by its advanced cultural behaviors.
There is general consensus that H. erectus evolved from H. habilis and Homo heidelbergensis evolved from H. erectus in Africa, eventually supplanting H. erectus populations in the Old World (Figure \(\PageIndex{0}\)).
Homo heidelbergensis
Some publications, e.g. Larsen 2014, refer to H. heidelbergensis as archaic Homo sapiens, but for our purposes, we will use the H. heidelbergensis designation. Otto Schoetensack found the first Homo heidelbergensis fossils in 1907 in Mauer, Germany. Since then H. heidelbergensis fossils have been found in Africa, Europe, and Asia. The date range for the species is 800 kya to 350 kya. Primitive traits include its large supraorbital tori, low frontal bone, sagittal keeling, and low frontal bone. Derived traits include separate supraorbital tori over each eye orbit, a more vertical posterior cranial vault, wide parietal bones in relation to the cranial base, and a larger cranial capacity than H. erectus (Becoming Human c2008). Additionally, they exhibit sexual dimorphism similar to that of modern humans.

Figure \(\PageIndex{3}\) - Homo heidelbergensis
There is regional variation in the morphology of H. heidelbergensis. European specimens found at Atapuerca, (Spain), Petralona (Greece), Seinheim (Germany), and Swancombe (England) show that they had compact bodies, which could have been a response to living in the cold climates of the north as it would help to conserve heat. Additionally, the cranium is a mosaic of H. erectus traits and derived traits. In Asia, data from sites such as Zhoukoudian, Jinniushan, and Dali (China) show a mix of H. erectus and H. sapiens traits; the latter includes large cranial capacity and thin braincase walls. African specimens from Kabwe (Zambia), Florisbad (South Africa), Laetoli (Tanzania), and Bodo (Ethiopia) also show a combination of H. erectus and H. sapiens traits. It shares the massive supraorbital tori and prominent occipital torus with H. erectus and thin cranial vault bones, less angulated occipital, and cranial base with H. sapiens.
H. heidelbergensis is the common ancestor of Homo neanderthalensis in Europe and Homo sapiens in Africa.
Homo neanderthalensis
Numerous Neanderthal fossils have been recovered since its discovery in 1856 in the Neander Valley, Germany. While the name of the person who found the first fossil is unknown it was described and named by William King. Neanderthals have been the speculation of scientists and the general public ever since. Some anthropologists classify Neanderthals as a subspecies of Homo sapiens, Homo sapiens neanderthalensis, while others interpret the morphological differences as significant enough to warrant classifying them as a different species, Homo neanderthalensis. In this essay, we will use the latter designation.
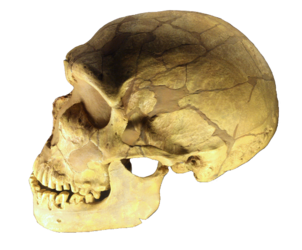
Figure \(\PageIndex{4}\) - Homo neanderthalensis
True Neanderthals first appear in the fossil record about 200 kya, with fossils exhibiting Neanderthal-like characteristics appearing as early as 400 kya. Recent research indicates that Neanderthals went extinct between 41 kya and 39 kya (Higham et al. 2014). Molecular research denotes that some Neanderthal DNA lives on in modern humans, approximately 2% of the DNA of “people who descend from Europeans, Asians, and other non-Africans is Neanderthal” (Callaway 2014). The Neanderthal genes are involved in fighting infections, dealing with ultraviolet radiation (Callaway 2014), and living at high altitudes (Callaway 2015). Neanderthal DNA has recently been linked with depression, obesity, and certain skin disorders, e.g., lesions caused by sun exposure (Callaway 2015).
Neanderthals are the only hominin to originally evolve in a glacial environment, leading to some characteristics adapted for the cold climate. Many scientists contend that the midface prognathism allowed for enlarged sinuses that functioned to warm and add moisture to the cold, dry air before entering the lungs. Small holes below the eye orbits, called the infraorbital foramina, are larger in the European Neanderthals that modern humans, suggesting that the blood vessels were larger, which allowed for more blood flow to the face. This would have helped keep the face warmer. They are relatively short and stocky, males averaging 5’ 5” and females 5’1”, with shorter appendages, both of which would help to conserve heat by providing less surface area from which to radiate heat. Their leg bones are thick and dense, suggesting that they frequently walked and ran, most likely in food procurement activities. Some postulate that some Neanderthals had pale skin that would have helped increase Vitamin D synthesis by increasing the amount of UV radiation to be absorbed by the body (O’Neil c1999-2014). The width of their body trunks and short tibias fit the predictions for cold weather adaptation as proposed by Christopher Ruff (Larsen 2013). The brain size of Neanderthals may also be related to cold weather. Averaging a cranial capacity between 1300 to 1400 cm3, they have the largest brains of all hominins, including H. sapiens. The size may be associated with increased metabolic efficiency in cold weather, which is similar to modern Inuit peoples today who have a larger brain size than other human populations (Jurmain et al. 2013).
The presence of an occipital bun is one of the characteristics used to identify H. neanderthalensis specimens, although it should be noted that this characteristics persists in a small percentage of modern human populations. The occipital bun may have evolved to counterbalance Neanderthal’s heavy face when running; it prevents the head from making huge horizontal accelerations (NOVA 2002). The occipital bun also makes the skull elongated in comparison to H. sapiens. They had heavy brow ridges like those of H. heidelbergensis. Neanderthal skulls recovered from Amud and Tabun in Israel exhibit more H. sapiens-like cranial characteristics, including lack of an occipital bun, smaller eye orbits, tall and wide nasal openings, and smaller teeth (Larsen 2014).
Denisovans
In 2010, scientists announced the discovery and DNA analysis of a finger bone and two teeth found in Denisova Cave, Siberia (Reich et al. 2010). The artifacts were recovered from a deposit dated to 50 kya to 30 kya. Data suggest that the remains were from an individual who shared a common origin with Neanderthals, but was not a Neanderthal, nor was it a modern human. The Denisovan individual(s) share 4-6% of its genetic material with modern peoples living in New Guinea, Bougainville Islands, and China. Further studies (Cooper and Stringer 2013) indicate that the Denisovans crossed Eurasia and interbred with modern humans, but genetically the Denisovans were more closely related to Neanderthals then modern humans (Meyer et al. 2012). While further finds will shed more light on the Denisovans, it is clear that there was more genetic variability during the Pleistocene then previously thought (Larsen 2014), but that gene flow between the various populations had implications for the emergence of modern humans (Pääbo 2015).
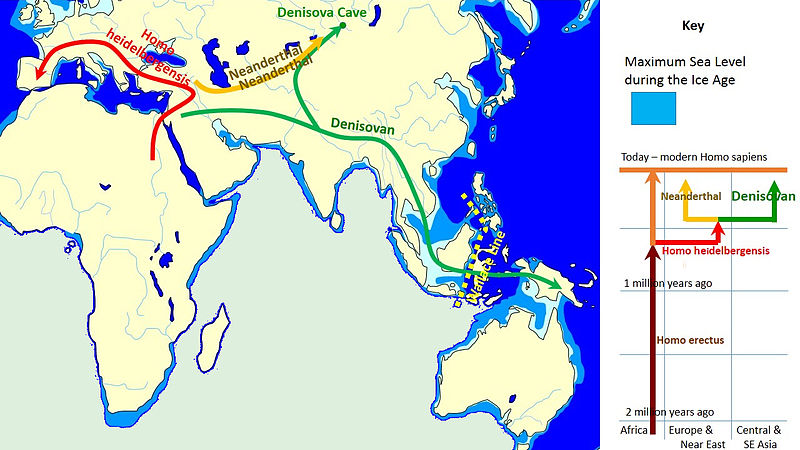
Figure \(\PageIndex{5}\) - Spread and Evolution of Denisovans
Homo floresiensis
In 2003, Peter Brown and Michael Morwood led a team that discovered unusual fossils in Liang Bua cave, Flores Island, Indonesia. The small stature of the individual led to the naming of a new species, Homo floresiensis. Since then twelve individuals have been found ranging in time from 74 kya to 17 kya. H. floresiensis had a small brain and stood only about 3.5’ tall as an adult. They had receding foreheads, no chins, shrugged-forward shoulders, and large feet relative to their short legs. They share some characteristics with H. sapiens, including smaller dentition, separate brow ridges, and a non-prognathic face.

Figure \(\PageIndex{6}\) - Homo floresiensis
Several hypotheses have been proposed to explain the appearance of H. floresiensis. One hypothesis suggests that we are seeing island dwarfism, which is an evolutionary process resulting from long-term isolation on a small island. Limited resources and lack of predators selects for smaller bodied individuals who need fewer resources then large-bodied individuals. Another hypothesis claims that H. floresiensisis not a separate species, but H. sapiens exhibiting microcephaly or some other developmental deficiency such as hyperthyroid cretinism (Oxnard et al. 2010) because the “…cranial features…are within the modern range of variation seen in living populations from the larger region [of Indonesia]” (Larsen 2014: 409), lending support to the second hypothesis. How H. floresiensis fits into the evolutionary picture is unclear based on the current data. An overview of the various hypotheses about its evolutionary relationships in the article, Homo floresiensis: Making Sense of the Small-Bodied Hominin Fossils from Flores.
Homo sapiens
At present, only one hominin species inhabits planet Earth, Homo sapiens. Morphologically, H. sapiens characteristics include the presence of a chin (or mental prominence), a large brain, flat face, rounded or globular cranium, and a continuous, reduced brow ridge. Its bones are gracile in comparison to earlier hominins, although the earliest H. sapiens are more robust than modern populations and shows none of the cold weather adaptations of Neanderthals. The more gracile nature lends credence to the hypothesis that modern humans evolved first in Africa. The leaner body proportions are more adaptive to the tropical African environments as there would be more body surface area to radiate heat. DNA evidence also supports an African origin. Modern African populations have more genetic diversity then any other modern human population, inferring that they have been evolving longer (Becoming Human c2008).
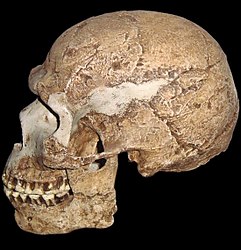
Figure \(\PageIndex{7}\) - Homo sapiens
The oldest H. sapiens fossils were found in Africa at Omo Kibish, Ethiopia, dated to 195 kya. H. sapiens fossils found at Herto, Ethiopia (160 kya–154 kya), Klasies River Mouth and Border Cave, South Africa (120 kya – 89 kya), Skhūl, Israel (130 kya–100 kya), and Qafzeh, Isreal (120 kya–92 kya), adds further support. Sites in Asia, e.g., Zhoukoudian and Tianyuan in China and Niah Cave, Borneo, Indonesia date to 40 kya or younger and in Europe dating no earlier then 31 kya (Mladeč, Czech Republic). Modern humans made it to Australia by 55 kya; however, no human remains have been found on the continent dating earlier than 30 kya at Lake Mungo. This data lends little support for the Multiregional Hypothesis, a model developed by Milford Wolpoff proposed that modern H. sapiens evolved in separate places in the Old World from local archaic H. sapiens populations. This model suggested that gene flow between the populations led to the genetic similarity of modern humans. Most anthropologists support the model of an African origin. There are two variants of this model. The Out of Africa model, proposed by Chris Stringer, claims that modern humans arose in Africa and spread to Europe after 50 kya, replacing non-H. sapiens population with no gene flow. Fred Smith and Erik Trinkhaus proposed the Assimilation model that also claimed modern human first arose in Africa and spread to Europe and Asia. The primary difference between the Out of Africa and Assimilation models is that the latter does claim gene flow between H. sapiens and H. neanderthalensis. Taking into account the recent DNA analyses of Neanderthals discussed above, multiple lines of evidence support the Assimilation model.
The case of the disappearing Neanderthals
We know that some Neanderthal genes are in the genome of modern humans, so in essence, some part of the Neanderthals survives today. However, there are no Neanderthals walking around in the modern world. So, what happened to the Neanderthals? Scientists suggest that H. sapiens out-competed Neanderthals with their more diverse diet and “sophisticated and cognitive abilities” (Becoming Human c2008). These traits allowed H. sapiens to readily adapt to rapidly changing climatic conditions during the Upper Paleolithic.
A recent hypothesis suggests that Neanderthals died out during a period of powerful volcanic activity in western Eurasia. Excavations at Mezmaiskaya Cave in the Caucasus Mountains in southern Russia have recovered a plethora of Neanderthal bones, stone tools, and remains of prey animals. The Neanderthal remains and artifacts appear in strata above a layer of volcanic ash and below a second layer of volcanic ash. No Neanderthal bones or artifacts have been found above the second level, suggesting that Neanderthals no longer occupied the area. Both of the ash layers contain levels of pollen associated with cooler, drier climates. “The ash layers correspond chronologically to what is known as the Campanian Ignimbrite super-eruption, which occurred around 40,000 years ago in modern day Italy, and a smaller eruption thought to have occurred around the same time in the Caucasus Mountains” (University of Chicago Press Journals 2010). The ensuing volcanic winter caused a dramatic climate shift that led to the demise of the Neanderthals. This data correlates with Higham et al.’s (2014) research that claims Neanderthals went extinct between 41 kya and 39 kya.
References
- Becoming Human. c2008. The human lineage through time. Institute of Human Origins [Internet] [cited 2015 Aug 3]. Available from: http://www.becominghuman.org/node/human-lineage-through-time
- Callaway E. 2014. Modern human genomes reveal our inner Neanderthal. Nature [Internet] [cited 2015 Aug 21]. Available from: http://www.nature.com/news/modern-hu...erthal-1.14615
- Callaway E. 2015. Neanderthals had outsize effect on human biology. Nature [Internet] [cited 2015 Aug 21]; 523 (7562): 512-513. Available from: http://www.nature.com.offcampus.lib....iology-1.18086. doi: 10.1038/523512a
- Cooper A, Stringer CB. 2013. Did the Denisovans cross Wallace’s Line? Science [Internet] [cited 2015 Aug 21}; 342 (6156): 321-323. Available from: http://www.sciencemag.org.offcampus....d-963a99b700ea. doi: 10.1126/science.1244869.
- Dembo M, Matzke NJ, Mooers AØ, Collard M. 2015. Bayesian analysis of a morphological supermatrix sheds light on controversial hominin relationships. Proc R Soc B [Internet] [cited 2015 Aug 20]; 282(1812). Available from: http://rspb.royalsocietypublishing.o...0943.e-letters. doi: 10.1098/rspb.2015.0943.
- eFossils [Internet] [cited 2015 Aug 10]. Department of Anthropology, The University of Texas at Austin. Available from: http://efossils.org/
- Higham T, Douka K, Wood R et al. 2014. The timing and spatiotemporal patterning of Neanderthal disappearance. Nature [Internet] [cited 2015 Aug 21]; 512 (7514): 306-309. Available from: http://www.nature.com.offcampus.lib....ture13621.html. doi:10.1038/nature13621
- Jurmain R, Kilgore L. Trevathan W. 2013 Essentials in physical anthropology. Belmont (CA): Wadsworth Cengage Learning.
- Larsen, CS. 2014. Our origins: discovering physical anthropology. New York (NY): W. W. Norton & Company, Inc.
- Lice and human evolution [Internet]. 2011 Feb 6. NOVAscienceNow. [cited 2015 Aug 20]. Available from: www.pbs.org/wgbh/nova/evolution/lice.html
- Meyer M, Kircher M, Gansauge MT, et al. 2012. A high-coverage genome sequence from an archaic Denisovan individual. Science [Internet] [cited 2015 Aug 21]; 338 (6104): 222-226. Available from: http://www.sciencemag.org.offcampus..../6104/222.full. doi: 10.1126/science.1224344.
- NOVA [Internet]. 2002 Jan 22. Neanderthals on trial. PBS [cited 2015 Aug 21]. Available from: http://www.pbs.org/wgbh/nova/transcr...nderthals.html
- O’Neil D. c1999-2012. Early human evolution: a survey of the biological and cultural evolution of Homo habilis and Homo erectus. Behavioral Sciences Department, Palomar College [Internet] [cited 2015 Aug 16]. Available from: anthro.palomar.edu/homo/Default.htm
- O’Neil D. c1999-2014. Evolution of modern humans: a survey of the biological and cultural evolution of archaic and modern Homo sapiens. Behavioral Sciences Department, Palomar College [Internet] [cited 2015 Aug 16]. Available from: anthro.palomar.edu/homo2/default.htm
- Oxnard C, Obendorf PJ, Kefford BJ. 2010. Post-cranial skeletons of hyperthyroid cretins show a similar anatomical mosaic as Homo floesiensis. PLoS One [Internet] [cited 2015 Aug 21]; 5 (9). Available from: http://www.sciencedaily.com/releases...0928025514.htm. doi: 10.1371/journal.pone.0013018.
- Pääbo S. 2015. The diverse origins of the human gene pool. Nat Rev Genet [Internet] [cited 2015 Aug 21]; 16: 313-314. Available from: http://www.nature.com.offcampus.lib....l/nrg3954.html. doi:10.1038/nrg3954
- Reich D, Green RE, Kircher M, et al. 2010. Genetic history of an archaic hominin group from Denisova Cave in Siberia. Nature [Internet] [cited 2015 Aug 21]; 468 (7327). Available from: http://www.nature.com/nature/journal...ture09710.html
- Smithsonian Institution [Internet]. 2015 Aug 4. What does it mean to be human? [cited 2015 Aug 11]. Available from: http://humanorigins.si.edu/evidence/human-fossils/species
- University of Chicago Press Journals [Internet]. Volcanoes wiped out Neanderthals, new study suggests. ScienceDaily 2010 Oct 7 [cited 2015 Aug 21]. Available from: http://www.sciencedaily.com/releases...1006094057.htm
- Villmoare B, Kimbel WH, Seyoum C, Campisano CJ, DiMaggio EN, Rowan J, Braun DR, Arrowsmith JR, Reed KE. 2015. Early Homo at 2.8 MA from Ledi-Geraru, Afar, Ethiopia. Science [Internet]. [cited 2015 Aug 16]; 347(6228): 1352-1355. Available from: http://www.sciencemag.org.offcampus....6228/1352.full. doi: 10.1126/science.aaa1343