
2.5: Evolutionary Theory (Part 3)
- Last updated
- Save as PDF
- Page ID
- 6654

Science doesn't stop with a founder. Creationists blame Darwin for evolutionary theory, but most biologists wouldn't call themselves “Darwinists”, any more than most physicists wouldn't call themselves “Newtonists.” Natural selection is just one factor of evolutionary theory. People know much more about evolution today than they did during Darwin's time. That doesn't mean that Darwin was wrong, just that science has progressed.
Darwin often gets blamed for Social Darwinism, the political ideology that extends "survival of the fittest" to justify exploiting the poor, but Darwin didn't come up with this phrase nor did he apply Natural Selection to human society. Social Darwinism was invented and promoted by others such as Herbert Spencer, Thomas Huxley, and Francis Galton. Charles Darwin objected to the application of his biological model to human social structure, and he definitely would have objected to the "Darwin Project" battle royale game, and "Darwin Awards" given out in his name.
So far, our overly broad unifying theories haven't been worth much more than interesting metaphors, so please try to separate nature from nurture: biology from culture.
Gregor Mendel
Even though neither Darwin nor Mendel knew about how heredity worked at the cellular level, it's almost impossible to talk about the consequences of their work without referring to what we know now. So we will introduce a few terms in this section that are anachronistic, and we'll wait to explain them in depth until the section on cellular biology.
The genius of Mendel is how he used mathematics to show how inheritance worked.
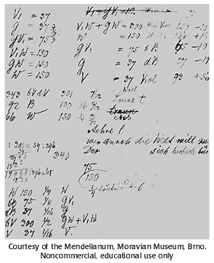
Figure \(\PageIndex{22}\) - Mendel counted peas
Dominance and Recessiveness
Remember that Gregor Mendel (1822-1884) didn't know about DNA when he did his experiments, he didn't see meiosis in the microscope, he wasn't directly involved in the debates over evolution, but he found one of the sources of variation that Darwin's theory of natural selection relies on, and he discovered two important principles that are the foundation of genetics: The Principle of Segregation and The Principle of Independent Assortment. Darwin knew that variation was crucial to his theory, but he didn't know the source of variation.
The pea plant has variation. Some seeds are smooth, some wrinkled; some yellow, some green. Some pods are inflated, some constricted; some green, some yellow. Some flowers are purple, some white; some along the stem, some at the top. Some stems are tall, some are short. Mendel was careful to exclude other kinds of variation: how some plants are eaten by snails, some don't get enough water, some too much sun, some are cooked in soup, some peas are overcooked, some shot through straws. Mendel ignored all these things that happen to peas and only paid attention to this first set of variations, the either/or inherent characteristics that can be seen.
Mendel was rediscovered around 1900. Theories of inheritance at the time of Mendel focused on blending, for example, one parent with extremely dark skin and one parent with extremely light skin have a child who is neither very light, nor very dark, but a color that is in between the extremes. But when Mendel bred purple flowers with white flowers, he got only purple flowers, and then when he bred those purple flowers together, in the next generation he got mostly purple but some white ones. The white flower trait disappeared and then came back. The purple color dominated the white one, but the recessive white color was not gone for ever, it came back in a later generation. If you cross a purple flower with a white flower, Darwin would have expected a whitish-purple flower. What happened to the blending?
If you don't remember from your high-school biology class, here are some basics that we got from Mendel:
- Children resemble their parents.
- Genes come in pairs.
- Genes don't blend.
- Some genes are dominant.
The Principle of Segregation
Good science can come from unlikely sources. Mendel would momentarily escape from the duties of a monk in a cold monastery.
Mendel frequently took sanctuary in the little two-room building nestled into a corner of the monastery courtyard right up against the brewery next door. It gave him not only blessed warmth but also the space to engage in his scientific pursuits -- which would, he believed, prove important enough in time to earn him a place in the annals of horticulture. He had filled the glasshouse's long tables with pots of pea plants, each carefully labeled as to seed source and variety. His immediate goal was to breed these peas, thirty-four different seed types in all, after allowing them to self-fertilize for two full years. In the speeded-up growing seasons of the glasshouse, two years of growing meant perhaps six full generations -- enough to assure Mendel that the seeds were indeed what they appeared to be. [Marantz Henig 2000:14]
Mendel isolated and bred different pea plants together and observed the characteristics of their offspring, and what really amazes me is that he counted them--and looking at the numbers he noticed patterns, and used simple math to work out the ratios of different traits.
First, he crossed true breeding plants (P) to get heterozygous plants, called hybrids (F1), and then he crossed those hybrids with each other (F1 x F1 = F2) and counted:
Expt 1: Form of seed. –– From 253 hybrids 7,324 seeds were obtained in the second trial year. Among them were 5,474 round or roundish ones and 1,850 angular wrinkled ones. Therefrom the ratio 2.96:1 is deduced.
Expt 2: Color of albumen. –– 258 plants yielded 8,023 seeds, 6,022 yellow, and 2,001 green; their ratio, therefore, is as 3.01:1.
[...]
Expt. 3: Color of the seed–coats. –– Among 929 plants, 705 bore violet–red flowers and gray–brown seed–coats; 224 had white flowers and white seed–coats, giving the proportion 3.15:1.
Expt. 4: Form of pods. –– Of 1,181 plants, 882 had them simply inflated, and in 299 they were constricted. Resulting ratio, 2.95:1.
[...]
Expt. 5: Color of the unripe pods. –– The number of trial plants was 580, of which 428 had green pods and 152 yellow ones. Consequently these stand in the ratio of 2.82:1.
Expt. 6: Position of flowers. –– Among 858 cases 651 had inflorescences axial and 207 terminal. Ratio, 3.14:1.
Expt. 7: Length of stem. –– Out of 1,064 plants, in 787 cases the stem was long, and in 277 short. Hence a mutual ratio of 2.84:1. In this experiment the dwarfed plants were carefully lifted and transferred to a special bed. This precaution was necessary, as otherwise they would have perished through being overgrown by their tall relatives. Even in their quite young state they can be easily picked out by their compact growth and thick
dark–green foliage. [Mendel 1865]
Notice that the ratios of the F2 generation work out to about 3:1 which means that three plants have the dominant phenotype for every one plant that has the recessive phenotype. Mendel showed that each trait (seed color, seed shape, pod shape, pod color, flower color, flower position, stem length) is determined by a pair of characters, and they get them from their parents, one from the pollen cell and one from the egg cell, which come together to form the embryo. When the pollen and egg cells are made, these two characters are "segregated" so each egg and pollen cell has only one character. In genetics we now call these traits, genes, and the pair of characters is called a pair of alleles. From cellular biology, we now know that the segregation of alleles during the production of eggs and sperm is called meiosis. We'll come back to this in the sections on genetics and cellular biology.
The Principle of Independent Assortment
If you take true breeding plants with two different traits, like form of seed and color of seed-coat, cross them together, you first get all of the dominant trait. Then if you cross those new versions again, you get some interesting numbers 9:3:3:1 The numbers reveal that there's no connection between the traits; the traits are independently assorted. We can now explain this with cellular biology because the two traits are on different chromosomes.
Punnett squares
A Punnett square is a grid or matrix that represents the outcomes of different combinations. They are often presented as proofs of Mendel's Principle of Segregation and Principle of Independent Assortment, but Punnett squares came after Mendel, and I think it's important to understand the steps Mendel went through in his research: empirical observations of pea plant variations, breeding true-breeding plants, crossing specific traits, getting weird results, counting them, working out simple ratios, explaining the ratios as biological Principles as to how the peas (and all life, including humans) reproduce and transmit the information using traits from parent to offspring. Punnett squares are graphic representations of sexual reproduction: all the possible sperm are one axis, all the possible eggs on the other, and in the middle are all the possible combinations of fertilization – the individual zygotes (fertilized egg) who develop into fetuses, babies, and then adults. About a hundred years after Mendel's experiment we got to look in a microscope to confirm Mendel's mathematics and we continue to explore Mendelian traits in humans.
Exercise \(\PageIndex{1}\)
Read: "MENDELIAN LAWS APPLY TO HUMAN BEINGS"
Here is an example using Tay-Sachs disease. The * HEXA gene on chromosome 15 makes part of an enzyme that is important for maintaining your central nervous system. If you have one or two normal alleles, you're OK, but if both your alleles have a Tay-Sachs mutation, then you'll have different neurological problems usually starting as an infant. If you are a genetic counselor and a couple comes to you planning to have kids, and they are both carriers (heterozygotes), you want to be able to tell them what is the chance their baby will have Tay-Sachs. If we assign symbols to alleles, "t" = a Tay-Sachs mutation, and "T" = normal HEXA allele, then we can diagram the possible outcomes of fertilization.
| T | t | |
| T | TT | Tt |
| t | Tt | tt |
Statistically, 25% of their children will be normal (TT), 50% of their children will be carriers (Tt), and 25% of their children will be born with Tay-Sachs (tt). This principle works with most recessive diseases.
Figure \(\PageIndex{23}\)
Mendelian Traits Laboratory
Your phenotype results from the interaction of your genotype and the environment. Most traits are polygenic, meaning several genes contribute to how they are expressed. Even though your genes guide your development, the environment where you grow up influences how those genes are expressed. The combination of polygenic and environmental influence leads to an amazing variety of individuals. However, humans have a small number traits that are readily observable because they are: 1) Mendelian (determined by a single gene) so their expression is on/off, 2) tend not to be effected by the environment, 3) have high enough allele frequencies that someone in the class probably expresses them, and 4) are visible without genetic testing.
For each of the traits described below, your assignment is to: 1) record your phenotype, 2) assign letters to represent the alleles of the gene, and 3) list your possible genotypes. Your phenotype is best described in a complete sentence: "I can ____.", "I cannot ___."; "I have ____.", "I don't have ____." Include only the trait that you express. Mendelian genetics assigns letters or combinations of letters to represent alleles. For two-allele genes, often a single letter is used, the capital letter for the dominant allele and the lowercase letter for the recessive allele. For a two-allele gene, you can have three possible genotypes: heterozygous, homozygous dominant, and homozygous recessive. Depending on your phenotype, you may have more than one possible genotype that leads to the phenotype that you observed. [text adapted from Mendelian Traits Laboratory; missing source]
Example \(\PageIndex{1}\): PTC tasting
PTC, or phenylthiocarbamide, is a human-made chemical. While the majority of people find PTC to have a bitter taste, many find this substance tasteless. To discover your phenotype, chew a strip of filter paper that has been soaked in a concentrated solution of PTC. The ability to taste PTC is inherited as a dominant. There are some studies that compare tasting PTC to tasting broccoli. There are several genes and environmental influence involved but "PTC tasting is largely determined by a single gene, TAS2R8, with two common alleles, and the allele for tasting is mostly dominant over the allele for non-tasting" (McDonald 2012).
You would write:
PTC Tasting
My phenotype: Broccoli tastes bitter to me, and I can taste PTC.
Alleles: B= the allele that codes for tasting PTC; b= the allele that codes for not tasting PTC
My possible genotypes: BB or Bb
or depending on your phenotype you might write:
PTC Tasting
My phenotype: Broccoli tastes good to me, and I can't taste PTC.
Alleles: B= the allele that codes for tasting PTC; b= the allele that codes for not tasting PTC
My possible genotypes: bb only
Earwax
Earwax, or cerumen, occurs in two basic forms. The dry form is gray and brittle while the wet form is brown and sticky. The dry form is inherited as a recessive. The gene is located on *chromosome 16

Figure \(\PageIndex{24}\) - "Mimikaki -- Japanese Ear Picks" © 2014 Dr. Timothy C. Hain

Figure \(\PageIndex{25}\) - "Cotton swab" by Aney. (CC BY-SA 3.0)
Mid-Phalanx Hair
Look at the middle segment (phalanx) of your fingers. Note the presence or absence of hair. Complete absence of hair reflects a homozygous recessive genotype. Note that some types of work may wear the hair away.
Lactase Persistence
The ability to digest milk as an adult is a dominant trait. This is good example of biocultural evolution: a biological trait intertwined with cultural factors (how people produce food).
* more info at Wikipedia
Relative Finger Length
Lay your right hand on a piece of lined paper with the fingers perpendicular to the lines. Note the relative lengths of the second (index) and fourth (ring) fingers. There are three possible situations: the second finger is longer than the fourth, the second finger is shorter than the fourth, or the second finger is the same length as the fourth.
Individuals who are homozygous for the allele for short index finger have a shorter index finger. Individuals who are homozygous for the allele for long index fingers have longer index finger. Individuals whose index and ring fingers are equal length are heterozygous. However, this is not a true Mendelian trait because the expressions of the two alleles in the heterozygous individual show that this trait is influenced by at least one other gene on the 23rd chromosome. Heterozygous males express an index finger that is equal to or shorter than the ring finger. But heterozygous females will have an index finger that is equal or longer than the ring finger.
Exercise \(\PageIndex{2}\): IS YOUR RING FINGER LONGER THAN YOUR INDEX FINGER?

Figure \(\PageIndex{26}\)5 -"Index finger longer than ring finger" by Arnie Schoenberg, adapted from "Gaye Holud and Mehendi" by Russell eee from Wikimedia Commons (CC BY-SA 4.0)
If so, you're up to three times more likely to develop arthritis of the knee than women whose ring fingers are the same length as or shorter than their pointers. The British scientists who discovered the link speculate that it may have to do with hormones. This finger pattern is more common in women with low estrogen levels. Another clue: Men are more likely to have longer ring fingers, but for them, there's less of an arthritis link. [Good Housekeeping 2008:37]
* Article on cave art and relative finger length
ABO Blood type
Read Dennis O'Neil on Human Blood
Remember not to confuse the ABO blood type, a gene located on the ninth chromosome, with the Rhesus factor (Rh+, Rh-) a gene located on the first chromosome.
Figure \(\PageIndex{27}\) public domain
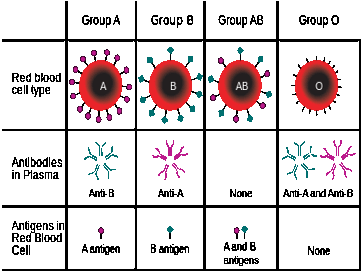
Figure \(\PageIndex{28}\) - en.Wikipedia.org/wiki/ABO_blood_group_system
Try to shift your thinking from Dominant means good/strong/prevalent, and Recessive means bad/weak/rare. Recessive just means it takes two alleles to expressive the trait. Dominant means you can have 2, or just 1 allele to express the trait. Don't conflate dominance with fitness. Dominance has to do with Mendel and inheritance, fitness is from Darwin's theory of natural selection.
Exercise \(\PageIndex{3}\)
Review Dennis O'Neil's Mendelian Genetics
*Skim Mendel, Gregor. 1865. "Experiments in Plant Hybridization"
*Play around with the Online Mendelian Inheritance in Man database. Try typing in the name of a disease, or body part into the search engine, and follow the link.
Population Genetics
By the 1920s, a new definition of evolution became popular: "a change in allele frequency". We're looking at how many people have a trait in one generation and how it might change in the next generation.
To translate a Punnett square into math, just replace the sperm and eggs with variables that represent allele frequencies, and you get a pretty simple algebraic equation, that can be used to study how populations change over time. It was discovered independently in 1908 by two scientists: Hardy and Weinberg.
| p | q | |
| p | pp | pq |
| q | pq |
A frequency is just another word for a percentage except we write it as a decimal e.g. ![]() = .5 is the same thing as saying 50% are p, or that half the alleles are p. This implies that the other half are q, because with a two-allele gene, the percentages have to add up to 100% (the frequencies add up to 1.00), so
= .5 is the same thing as saying 50% are p, or that half the alleles are p. This implies that the other half are q, because with a two-allele gene, the percentages have to add up to 100% (the frequencies add up to 1.00), so
\[p + q = 1\]
we can square both sides to get:
\[(p+q)^{2} = p^{2} + 2pq + q^{2} = 1\]
Now compare this to the Punnett square: pp is p2, qq is q2, and the two pq are 2pq. The p2 and q2represents the frequencies of the homozygotes and 2pq represents the frequency of the heterozygote. So you can take a population, count the total alleles, and count the phenotypes of the individuals and compare them to see if anything looks weird. Mathematically, you expect the numbers to work out, and when they don't, you know some kind of evolution is occurring.
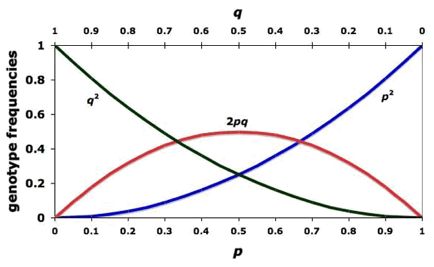
Figure \(\PageIndex{29}\) - How the allele frequencies are supposed to work without evolution; the null hypothesis
Exercise \(\PageIndex{4}\)
Read the Dennis O'Neil Intro to the Hardy-Weinberg Equation.
*and here is a longer introduction to Population Genetics, if the math freaks you out, just skim to later sections.
The Modern Evolutionary Synthesis
By the 1920s, evolutionary theory had synthesized the macroevolution of Darwin, with the microevolution of Mendel and population genetics, and came up a short list of factors that cause evolution. They tested populations with the Hardy-Weinberg equation, and when they failed to get a null hypothesis, they started trying to figure out which of the four forces of evolution caused the change in allele frequency: mutation, natural selection, genetic drift, and migration. The theories have been refined in the last hundred years, the four forces of evolution is still a useful way to think of evolution.
Vocabulary
- antibodies
- antigens
- codominant
- dominant
- fixity of species
- Great Chain of Being
- Hardy-Weinberg Equation
- macroevolution
- microevolution
- population
- the Principle of Independent Assortment
- the Principle of Segregation
- Punnett Square
- recessive