This appendix provides a very brief introduction to the EEG and ERPs (and necessarily oversimplifies some complex issues). It is meant to provide enough background for you to understand the big picture of the analysis procedures described in this book, but it is just a start.
The EEG
The neurons in your brain produce two main kinds of electrical potentials (voltages), called postsynaptic potentials and action potentials. We can’t ordinarily pick up action potentials from the scalp, but the postsynaptic potentials produced by individual neurons can propagate through the brain, skull, and scalp. If we put electrodes on the scalp, as shown in Figure A1.1, we can pick up the propagated postsynaptic potentials. These propagated voltages are what we call the electroencephalogram or EEG. This is a simplification; if you want to know the full story, I recommend Jackson and Bolger (2014) or Buzsáki et al. (2012).
The postsynaptic potentials produced by individual neurons sum together and spread widely before reaching the scalp, so any given electrode is picking up voltages generated by thousands or millions of neurons in a broad set of brain regions. As a result, a voltage picked up at a given electrode site may not represent activity coming from the cortex directly under the electrode.
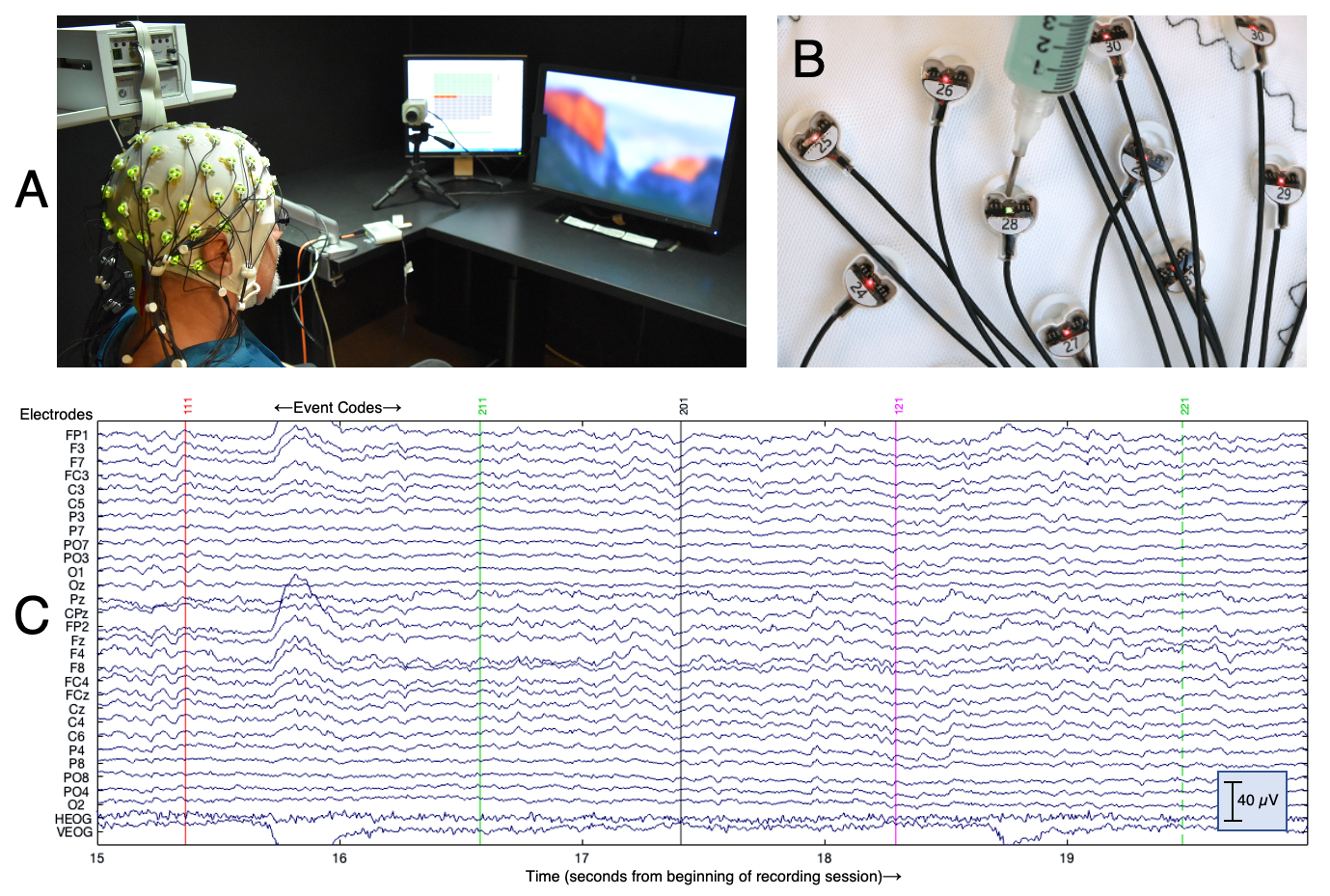
Figure A1.1. Typical EEG recording. (A) The subject is in the lab, wearing an electrode cap and facing the computer monitor where the stimuli will be presented. (B) Each electrode is a metal pellet encodes in plastic. A conductive gel is squirted into the electrode holder to make an electrical contact between the electrode pellet and the skin. (C) A 5-second period of EEG. Each waveform is the signal from one of the 30 electrodes that was used in this experiment. The X axis is time (in seconds), and the Y axis is voltage (in microvolts, µV). The colored vertical lines indicate event codes, which are numeric codes that mark the time and identity of the stimuli that were presented to the subject. The EEG signals are referenced to the right mastoid. HEOG (horizontal electrooculogram) is the potential between two electrodes placed just lateral to the eyes, and VEOG (vertical electrooculogram) is the potential between electrodes placed under and over the right eye.
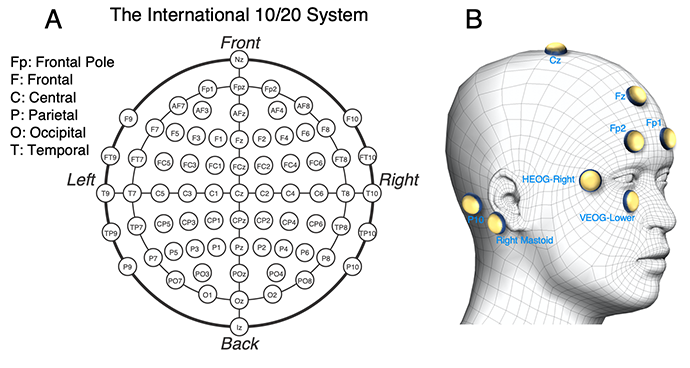
The EEG may be recorded from just a few electrodes or from as many as 256 electrodes, but most studies include between 16 and 64 electrodes. As shown in Figure A1.2, there is a widely used electrode naming system (the International 10/20 System) in which each electrode is given a 1-2 letters to indicate the general region of the head and a number to indicate the left-right position (or a “z” for the midline). For example, electrode P1 is over parietal cortex, just slightly to the right of the midline, and electrode FCz is on the midline, between the frontal and central regions.
Figure A1.2. (A) The International 10-20 system. In this system, each electrode is named with 1-2 letters indicating the general region of the scalp and a number indicating the left-right location. The numbers are odd for the left hemisphere, even for the right hemisphere, and get larger as we move away from the midline (which is labeled with a “z” to indicate “zero”). For example, Fp2 is over the frontal pole, just to the right of the midline, and Cz is over the central sulcus on the midline. (B) Additional common electrode sites. Electrodes are often placed over the left and right the mastoid processes (the bony protrusions just behind the earlobes) as reference electrodes. The horizontal electrooculogram (HEOG) is often recorded from electrodes just lateral to the left and right eyes to detect horizontal eye movements. The vertical electrooculogram (VEOG) is often recorded from an electrode just under the left or right eye (referenced to the corresponding Fp1 or Fp2 electrode over the eye) to detect blinks and vertical eye movements.
The Reference Electrode
A voltage is the potential for electrical charges to move from one place to another (e.g., from one terminal of a car battery to the other). As a result, the EEG is always a voltage between two electrodes, which we call the active electrode and the reference electrode. Typically, most or all of the active electrodes share the same reference electrode. Consider, for example, the EEG waveforms in Figure A1.1.C. Each waveform has a label, which indicates the active electrode for that waveform, and all of these channels except the bottom two used the same reference electrode, which was located on the mastoid process (the bony protrusion behind the ear, shown in Figure A1.2.B). This means that the waveform labeled F7 shows the electrical potential between the F7 electrode site and the mastoid. In theory, we would like a reference that is electrically neutral and does not impact the observed waveform. In practice, however, there is no neutral site, and the waveform for a given channel is equally impacted by activity arising from the active and reference electrodes.
The horizontal and vertical electrooculogram signals at the bottom of Figure A1.1.C (labeled HEOG and VEOG) use different reference electrodes. The HEOG signal uses HEOG-right as the active electrode (see Figure A1.2.B) and the mirror-image HEOG-left electrode as the reference. The VEOG signal uses the VEOG-lower electrode as the active and the FP2 electrode as the reference. These bipolar configurations make it easier to detect eye movements and blinks, which are large artifacts that propagate across the scalp.
Choosing the best reference electrode can be challenging. Fortunately, you can re-reference your data offline to any electrode (or combination of electrodes) that was in the original recording.
Artifact Rejection and Correction
Some portions of the EEG are contaminated by very large artifacts arising from sources such as eye blinks, eye movements, and muscle activity. In some studies, these epochs are excluded from all analyses, which is called artifact rejection. In other studies, a mathematical procedure is used to estimate the contribution of the artifacts and remove them from the epochs, which is called artifact correction. Many studies use correction for some artifacts and rejection for others.
Signal Averaging and Event-Related Potentials
The EEG is an incredibly complex signal that includes activity generated throughout the cortex from hundreds or even thousands of processes, all of which are summed together in our scalp electrodes. As a result, you can’t get a lot of information about specific neurocognitive processes from looking at the raw EEG. The raw EEG signal can tell you if someone is asleep, awake but zoned out, or highly alert and attentive, and you can sometimes see evidence of pathology (e.g., epileptic spikes). Signal processing methods must be applied to the EEG data to isolate specific processes. Perhaps the simplest of these methods is signal averaging, which we use to extract ERPs from the EEG.
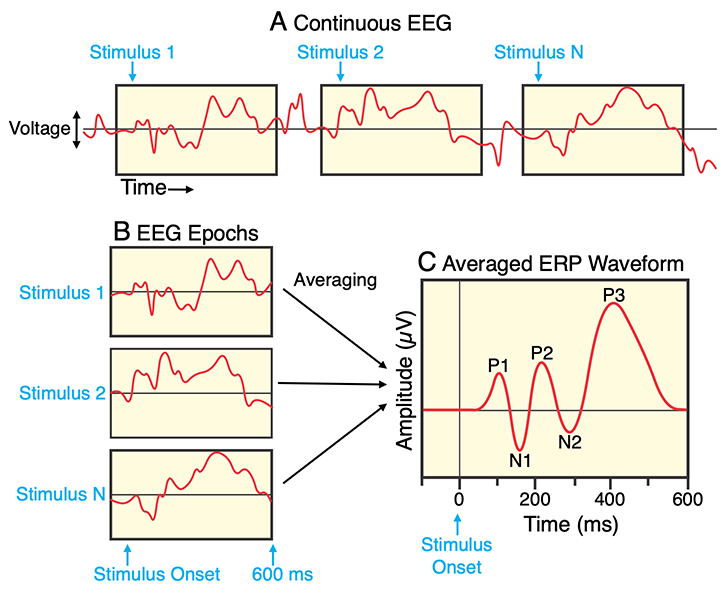
Signal averaging is illustrated in Figure A1.3. The goal of signal averaging is to isolate the electrical potentials (voltages) that are related to specific events (i.e., the event-related potentials). Signal averaging accomplishes this by assuming that an event such as a stimulus produces a similar waveform on each trial, and random noise is added to this waveform to produce the EEG that we record. If we average together the recorded EEG following many events, then the noise will “average out” and the signal will remain in the average. Figure A1.3.A illustrates the EEG from a single electrode. Whenever an event such as a stimulus or response occurs, an event code is inserted into the EEG data file to mark the time of the event. In this example, the events are stimuli (e.g., shapes flashed on a computer monitor).
Figure A1.3. The signal averaging process. The EEG is recorded as a continuous signal (A) while stimuli are presented. An event code marks the onset of each stimulus. An epoch of EEG data is extracted for each stimulus (B), beginning slightly before stimulus onset (e.g., 100 ms) and extending for a fixed period after stimulus onset (e.g., 600 ms). The single-trial epochs are lined up and then averaged together to create an averaged ERP waveform (C). That is, the averaged value at time t is the average voltage at time t across all N epochs. The X axis is time in thousandths of a second (milliseconds, ms), and the Y axis is voltage in millionths of a volt (microvolts, µV). In a typical experiment, there would be several different types of stimuli, and a separate averaged ERP waveform would be created for each stimulus type. We would also have a separate waveform at each electrode site.
For each event, we extract an epoch of EEG data surrounding that event. In this example, the epochs begin 100 ms before stimulus onset (giving us a 100-ms prestimulus baseline period) and extend until 600 ms after stimulus onset. As shown in Figure A1.3.B, all of the epochs for a given type of stimulus are then lined up with respect to time zero. The voltage at a given time for a given stimulus is assumed to be the consistent ERP at that time plus random noise. If we simply average all the epochs together, the noise will largely cancel out, and the averaged ERP waveform will mainly consist of the consistent ERP response (plus some residual noise because we always have a finite number of trials). The number of trials required to produce an acceptable average varies widely across experiments and depends on factors such as how large the ERP is and how much noise is in the EEG. Note that artifact rejection and/or correction are applied prior to averaging.
In ERP research, we use the term noise to refer to any uncontrolled variability in the EEG or in the averaged ERP waveform. We are assuming that the EEG consists of a consistent ERP signal on each trial plus random variability. Anything that causes the EEG to deviate from the consistent ERP signal is considered noise. If we repeated an experiment multiple times for a given participant (assuming no fatigue, no learning, etc.), any differences in the averaged ERP waveforms across repetitions would be a result of this noise. Some of this noise comes from electrical devices in the recording environment that is unintentionally picked up by the recording electrodes. Some of it comes from nonneural biological sources, such as blinks and muscle contractions. Some of it comes from brain activity that is unrelated to the stimuli. All else being equal, an averaged ERP waveform will have less residual noise if more trials are averaged together.
What Do We Mean by “Noise”?
In ERP research, we use the term noise to refer to any uncontrolled variability in the EEG or in the averaged ERP waveform. We are assuming that the EEG consists of a consistent ERP signal on each trial plus random variability. Anything that causes the EEG to deviate from the consistent ERP signal is considered noise. If we repeated an experiment multiple times for a given participant (assuming no fatigue, no learning, etc.), any differences in the averaged ERP waveforms across repetitions would be a result of this noise. Some of this noise comes from electrical devices in the recording environment that is unintentionally picked up by the recording electrodes. Some of it comes from nonneural biological sources, such as blinks and muscle contractions. Some of it comes from brain activity that is unrelated to the stimuli. All else being equal, an averaged ERP waveform will have less residual noise if more trials are averaged together.
ERP Peaks and Components
Figure A1.3.C shows a hypothetical averaged ERP waveform for a visual stimulus recorded at an occipital electrode site (with no noise). The X axis is time, with stimulus onset at time zero. The Y axis is amplitude. The prestimulus period is flat because there should be no consistent stimulus-related activity prior to the stimulus. The waveform begins to deviate from zero at approximately 50-60 ms, which is when visual information typically reaches visual cortex. There is no delay between the postsynaptic potentials generated in the brain and the voltage deflections that we see in an ERP waveform. As a result, the sequence of voltages over time in the ERP waveform corresponds to the sequence of sensory, cognitive, affective, and motor processes that follow the onset of a stimulus.
The ERP waveform consists of a set of positive-going and negative-going waves or peaks, which are related in a complicated way to a set of underlying components in the brain that reflect specific neurocognitive processes. We can’t directly see these components, but we try to draw inferences about them from the observed scalp ERP waveforms (see Chapter 2 in Luck, 2014, for a detailed discussion).
Most ERP components are named with a P or an N to indicate whether they are positive-going or negative-going, followed by a number to indicate the timing. For example, N1 is the first major negative wave and P3 is the third major positive wave. If the number is large (>10), it instead indicates the approximately latency of the peak in milliseconds (e.g., N170 for a negative-going wave that peaks at 170 ms). Sometimes, a component is given a name that reflects the conditions under which it is observed (e.g., the error-related negativity for a negative voltage that is present when the participant makes an incorrect response). Note that the relationship between an underlying component and the observed peaks and waves is complex, and the component names can sometimes be misleading. For example, the P1 elicited by a visual stimulus is completely unrelated to the P1 elicited by an auditory stimulus, but the P3 is the same for auditory and visual stimuli. A detailed discussion is provided in Chapters 2 and 3 of Luck (2014) and by Kappenman and Luck (2012).
To statistically analyze ERP data, researchers typically start by obtaining an amplitude or latency score from each individual participant’s averaged ERP waveform in each group or condition. Then these scores are entered into a statistical analysis that is much like the analysis of a behavioral variable such as response time. For example, you might find the peak latency (the time point when the voltage reaches its maximum value) for the P3 wave in each participant’s ERP waveform and then determine whether the latencies are significantly later in a patient group than in a control group using a t test.
References
Buzsáki, G., Anastassiou, C. A., & Koch, C. (2012). The origin of extracellular fields and currents—EEG, ECoG, LFP and spikes. Nature Reviews Neuroscience, 13, 407–420. https://doi.org/10.1038/nrn3241
Jackson, A. F., & Bolger, D. J. (2014). The neurophysiological bases of EEG and EEG measurement: A review for the rest of us. Psychophysiology, 51(11), 1061–1071. https://doi.org/10.1111/psyp.12283
Kappenman, E. S., & Luck, S. J. (2012). ERP components: The ups and downs of brainwave recordings. In S. J. Luck & E. S. Kappenman (Eds.), The Oxford Handbook of ERP Components (pp. 3–30). Oxford University Press.