
12.2: Triggering Drinking Behavior - Osmometric and Volumetric Thirst
- Page ID
- 137528
This page is a draft and under active development. Please forward any questions, comments, and/or feedback to the ASCCC OERI (oeri@asccc.org).
- Discuss the role of solutes in determining the movement of water in the body.
- Distinguish between intracellular and extracellular fluid.
- Compare and contrast osmometric and volumetric thirst.
Overview
When talking about thirst, we are talking not just about water in the body, but also the major solute (dissolved substance) we find in this water, salt or sodium chloride (NaCl). Regulating the body's water is necessarily impacted by the substances that are dissolved in that water. Thirst is a signal to the body that there has been a loss of fluid or, more specifically, that there is a fluid imbalance. About two-thirds of the body's water is intracellular, meaning within cells. The other third, the extracellular fluid, consists of interstitial fluid (the fluid bathing cells), cerebrospinal fluid (fluid in the ventricular system), and blood plasma (intravascular).
Because we effectively have two places where water is found (intracellular and extracellular), we have two systems for monitoring the body's fluid levels - and a variety of ways of inducing thirst, as well as different types of thirst. In addition, salt appetite is necessarily associated with one form of thirst as the need created by the fluid loss requires both salt and water to restore homeostasis. One system focuses on the levels of intracellular fluid and triggers osmometric thirst. The other monitors extracellular levels - more specifically, plasma or blood volume - and triggers volumetric thirst. Volumetric thirst is associated wth a neeed for both salt and water.
Intracellular Fluid Volume
Intracellular fluid volume is controlled by the concentration of solutes in the interstitial fluid (the fluid outside of cells). Under normal circumstances, the fluid outside and inside the cell is isotonic (the concentration of solutes is equal). If, however, the solute concentration is increased in the interstitial fluid (due to ingestion of solutes or loss of water), water will leave the cell by the process of osmosis. Osmosis is simply movement of water from an area of low solute concentration to an area of high concentration, water will move as needed to even out the solute concentration (Figure \(\PageIndex{1}\)). If the interstitial fluid (the fluid outside of the cell) is hypertonic (if it has a higher solute concentration), water will leave the cell. If the interstitial fluid is hypotonic, water will enter the cell. Both conditions may be dangerous - disrupting normal neuronal function. When there is a difference between the solute concentration of the interstitial fluid and the intracellular fluid, the movement of water by osmosis causes changes in intracellular volume.
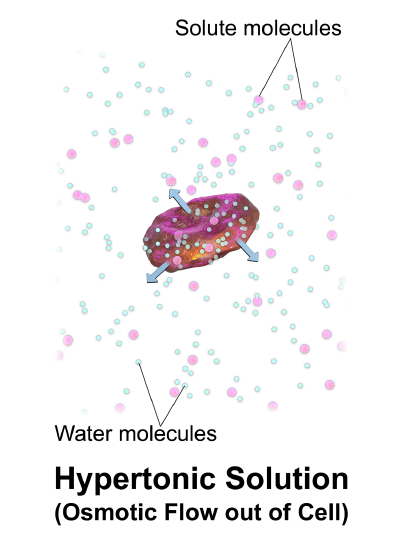
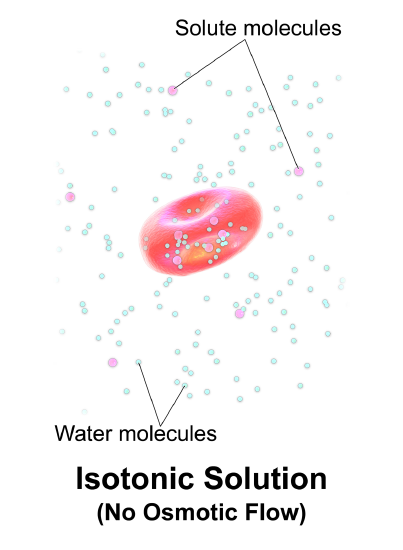

Generally, there is no need to regulate the volume of the interstitial fluid, although we do regulate its tonicity (solute concentration) through its effects on the intracellular fluid volume. More specifically, we monitor the movement of water.
Plasma Volume (Extracellular)
The other important and closely regulated fluid compartment is the blood plasma. If there is a loss of plasma volume (hypovolemia), this can impair the functioning of the heart. If increased, there may be a dangerous increase in blood pressure. If hypovolemia is severe enough the heart can no longer pump effectively. The vascular system can correct for loss of volume, but only within a limited range.
Two Variables, Two Regulatory Mechanisms
In order to maintain optimal fluid balance, the two variables are monitored:
- Movement of water (intracellular)
- Plasma volume (extracellular)
As there are two variables being monitored, there will be two different regulatory mechanisms that underlie thirst. The monitoring of fluid levels is tied to the monitoring of sodium levels. There are two mechanisms for dealing with a need for fluid and salt or sodium - we see both physiological and behavioral mechanisms for dealing with a need for water and salt:
- Physiologically - At the kidney, the excretion (loss) of water and salt can be modified. And alterations in heart rate and blood pressure can also compensate for losses.
- Behaviorally - ingest water and salt
What is the role of the kidneys and how is kidney activity regulated? Typically, we ingest much more salt and water than we need and what we don't need is excreted. Blood is essentially filtered by the kidneys. The functional units of the kidneys, the nephrons, extract fluid from the blood and carry it (collect it into) the ureter. From the ureter, urine passes to the urinary bladder where urine is stored.
As discussed in the prior section, two hormones are involved in the excretion of sodium and water by the kidney - both hormonal signals increase retention:
- Aldosterone is secreted by the adrenal cortex.
- Vasopressin (Anti Diuretic Hormone or ADH) is secreted by the posterior pituitary, but produced by the hypothalamus. The two names by which this hormone is known reflect two aspects of its effects, "vasopressin" is a reference to its ability to cause contraction of blood vessels while ADH reflects its role in preventing excretion.
What happens without vasopressin? No water is retained. Without vasopressin, diabetes insipidus develops, leading to excessive water loss and increased thirst. Incidentally, the term diabetes insipidus literally means "a tasteless passing through". The urine is so dilute, it has little taste. It is, as you might expect, treated with vasopressin - in the form of a nasal spray. The diabetes you more commonly hear of (due to a lack of or insensitivity to insulin) is technically diabetes mellitus. Diabetes, again, means "passing through". Mellitus means "sweet" as the urine of the diabetic would be sweet with unused glucose.
Osmometric (Osmotic) Thirst - Detecting Water Movement Out of Cells
So, we can drink to replenish lost fluid and the kidney's function can conserve or allow fluids to be lost. As we have two variables that we are monitoring, we have two different types of thirst. Osmometric thirst derives its name from the word osmosis. Osmometric thirst is triggered by loss of volume from the intracellular fluid stores. This form of thirst is produced when the solute concentration or tonicity of the interstitial compartment (the fluid outside of the cell) is increased, causing water to move out of the cell. Detectors, osmoreceptors, detect the loss of water. Interestingly, it is more than just the change in the solute concentration that triggers this form of thirst. It is the actual effect of this change on the water in the intracellular fluid. If water does not move out of the cell, drinking will not increase. If the concentration of interstitial solutes is increased with something that passes into the cell - so the extracellular and intracellular solute concentrations remain isotonic and there's no net movement of water - drinking will not be triggered. In other words, it isn't just changes in the solute concentration that are detected, it is the loss of water from the intracellular compartment that matters. An increase in fluid intake is only seen when there is an increase in interstitial solute concentration that results in the movement of water out of cells. The movement of water out of cells (cellular dehydration) triggers osmometric thirst.
Where is this change detected? Where are the osmoreceptors? We can find hints of this by looking at the effects of various solutes on thirst.
If urea is injected, the extracellular solute concentration is increased. Urea does pass into cells, but it only crosses the blood brain barrier slowly. In other words, there's no movement of water in the cells of the body, but as you’ve increased the solute concentration outside of the brain, there will be a net movement of water out of the brain. Not surprisingly, even a slight and temporary decrease in the intracellular fluid concentration of the brain results in thirst. It also makes sense that osmoreceptors would be found in the brain as the brain needs to be protected from changes and it is the brain that triggers behavior. A simpler (and less disturbing) way of demonstrating this is by injecting hypertonic saline directly into the brain. Osmoreceptors can be classified as central or peripheral osmoreceptors based on their location. The central receptors are primarily present in the anterior hypothalamus, including the organum vasculosum laminae terminalis (OVLT) and the subfornical organ (SFO) (Danziger & Zeidel, 2015; Xu et al., 2000; Muhsin & Mount, 2016). These central receptor cells have respond to osmotic changes and they are sensitive to angiotensin II (abbreviated as AII; Bichet, 2012; Benarroch, 2011). Some SFO/OVLT neurons also receive signals from peripheral arterial baroreceptors. Thus, SFO/OVLT neurons sense plasma osmolality, volume, and pressure to control thirst. These cells depolarize due to increased Na+ concentration, cell shrinkage, angiotensin II or negative suction pressure, and discharge neuronal spikes, which later initiate the sensation of thirst or vasopressin release or both (Muhsin & Mount, 2016; Gizowski & Bourque, 2018).
One of the central receptor organs, SFO, has two distinct types of neurons with opposing actions. A glutamatergic population (SFO-GLUT) that promotes thirst and sodium intake, and a GABAergic population (SFO-GABA) that inhibits thirst (Zimmerman, Leib, & Knight, 2017). The proper agonistic and antagonistic functioning of these cells maintain the optimum level of hydration. As the SFO has access to the systemic blood, it has a markedly different ependymal surface with a flattened appearance, lacks normal cilia count, and has tight junctions between adjacent cells. These features help block the diffusion of substances across the SFO parenchyma into the third ventricle. This peculiar position of the SFO cells is suitable for sensing both plasma and CSF components (Hiyama & Noda, 2016).
OVLT, the other central receptor, is close to the median preoptic nucleus. The peripheral receptors which are present within the upper gastrointestinal tract and portal venous system also detect changes in solute concentration and blood volume via a specific type of receptor (Danziger & Zeidel, 2015; Xu et al., 2000; Muhsin & Mount, 2016; Xu et al., 2000). They act as a supplementary center for osmoregulation in addition to the central osmoreceptors (Muhsin & Mount, 2016).
Volumetric (Hypovolemic) Thirst - Detecting Blood Loss
Volumetric thirst is triggered when there is a loss of volume from the extracellular fluid stores. Volumetric thirst is triggered by a decrease in blood volume referred to as hypovolemia. Hypovolemia is usually accompanied by a drop in blood pressure (hypotension). Hypovolemia and hypotension then ultimately lead to the production of angiotensin II. As discussed in the last section, a cascade of events is triggered when the kidneys detect a decrease in blood pressure and release the enzyme renin. This activates the renin-angiotensin-aldosterone system. Angiotensinogen, produced by the liver, is acted on by renin to produce angiotensin I, which is then converted into angiotensin II. Angiotensin II functions as a hormone and then causes the release of the hormone aldosterone by the adrenal cortex, resulting in increased sodium reabsorption, water retention, and an increase in blood pressure.
But how do we produce volumetric thirst experimentally? Obviously, we could remove blood. But removing a large volume of blood creates other problems. A better method is to inject a colloid. A colloid is a large molecule that can't cross cell membranes. By placing a colloid under the skin or in the abdominal cavity, extracellular fluid will move out of tissues. As the interstitial fluid decreases, fluid from the plasma fills the vacancy. As a consequence of the resulting drop in blood volume, vasopressin is released and the subject drinks. Early studies using this procedure documented the presence of both thirst (Fitzsimons, 1961) and sodium appetite (Stricker & Wolf, 1966) as indicated by ingestion of both. Later work (Stricker, 1981) explored the effects of colloid treatment on both water and saline drinking in greater detail. Stricker's work explored the effects of various manipulations on thirst and sodium appetite, revealing that sodium appetite accompanies thirst when dietary sodium is controlled. In other words, earlier studies had found that there was a delay in sodium appetite that may be accounted for by the excess of sodium that had previously been ingested. The separate timelines associated with thirst and sodium appetite indicate separate systems for detecting the needs for water and sodium.
Following extracellular fluid loss - loss of plasma volume - there is a need for both salt and water. A rat will now drink the saline solution that it normally rejects. This indicates that there is a mechanism for monitoring the body's sodium concentration, so that needs are detected and then mechanisms are employed to satisfy them. Unlike osmometric thirst, volumetric thirst is accompanied by a salt appetite.
Angiotensin II, the 3rd "step" when the renin-angiotensin-aldosterone system is activated, travels through the blood stream and exerts multiple effects.
- Angiotensin II acts at the adrenal cortex to stimulate aldosterone secretion.
- Angiotensin II acts at the posterior pituitary to stimulate vasopressin (ADH) secretion.
- Angiotensin II acts at the muscles of the the small arteries to cause contraction (increasing blood pressure).
- Angiotensin II triggers drinking and salt ingestion.
Although angiotensin II is thought to produce thirst and act on other brain sites, it does not cross the blood-brain barrier. Thus, in order to cause thirst and other effects, angiotensin II must act at a part of the brain that lacks this barrier. This is characteristic of the circumventricular organs (organs surrounding the ventricles), which includes the organum vasculosum laminae terminalis (OVLT) and the subfornical organ (SFO) mentioned earlier. The subfornical organ (SFO) appears to be the site at which angiotensin II acts to cause volumetric thirst.
Evidence for the role of the SFO in water ingestion:
- Very low doses of angiotensin II at the SFO cause drinking.
- Angiotensin II-induced drinking is blocked by blocking angiotensin II receptors at the SFO.
- Electrical stimulation of the SFO produces drinking.
Furthermore, neurons in the SFO increase their activity in response to angiotensin II, even when neural connections are cut - demonstrating that the neurons are responding to the AII.
The SFO has outputs to several brain regions. And these connections mediate the endocrine, autonomic, and behavioral responses associated with thirst. It is involved in a number of the effects of AII. How do the SFO and angiotensin II work together to produce all the effects associated with angiotensin II?
- Secretion of aldosterone by the kidney is a direct response to AII at the kidney. The other effects of AII are initiated at the brain.
- Endocrine outputs of the SFO communicate with the hypothalamic regions controlling release of vasopressin.
- Autonomic outputs of the SFO act on hypothalamic regions influencing the activity of the autonomic nervous system.
- Behavioral outputs of the SFO act on the basal forebrain. If these efferent connections of the SFO are lesioned, we no longer see angiotensin II-induced drinking.
So, there are receptors in the kidney that detect changes in blood flow. There is a second set of volumetric receptors in the heart. These are baroreceptors, they are stretch receptors that detect the amount of blood in the heart's atria (the part of the heart that is receiving blood from the veins).
Normal Drinking
Just as we typically eat long before a physiological need for fuel would trigger hunger, we drink before change in the body's fluid stores would be detected. As with eating, we have learned elements of our drinking behavior. When do we habitually drink? Normally, we drink with meals. While eating does produce a need for fluid, we typically drink before that need would normally be detected. Food in the digestive system causes water to be diverted to these areas and absorbed food increases the solute concentration of the plasma and produces osmometric thirst.
Why do animals drink before a need is detected? Animals will actually learn to drink more if their diet is changed such that they need to drink more. With a high protein diet, more water is needed. Animals will learn to drink more with a meal in anticipation of the need.
What signals or causes drinking with a meal? The movement of water into the digestive system produces hypovolemia. The hypovolemia causes the kidneys to secrete renin and AII levels are increased. During a normal meal the levels of renin actually double. If we then block AII production, we'll see a decrease in drinking with a meal.
Summary
In order to maintain an appropriate fluid balance, intracellular and extracellular fluid levels are monitored. Intracellular or osmometric thirst is triggered when water moves out of cells. In contrast, extracellular thirst and an appetite for salt is triggered when the loss of blood volume creates a need for salt and water.
Attributions
"Osmometric Thirst - Detecting Water Movement Out of Cells" adapted from Koshy, R. & Jamil, R. (2021). Physiology, Osmoreceptors. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2022 Jan-. Available from: https://www.ncbi.nlm.nih.gov/books/NBK557510/ (CC BY)
References
Leib, D. E., Zimmerman, C. A., & Knight, Z. A. (2016). Thirst. Current biology : CB, 26(24), R1260–R1265. https://doi.org/10.1016/j.cub.2016.11.019
Stricker E. M. (1981). Thirst and sodium appetite after colloid treatment in rats. Journal of comparative and physiological psychology, 95(1), 1–25. https://doi.org/10.1037/h0077764