
8.5: Touch and Pain
- Page ID
- 10645

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Introduction

Sensation
Cutaneous Senses of the Skin Connect the Brain to the Body and the Outside World
Different Receptor Types Are Sensitive to Specific Stimuli
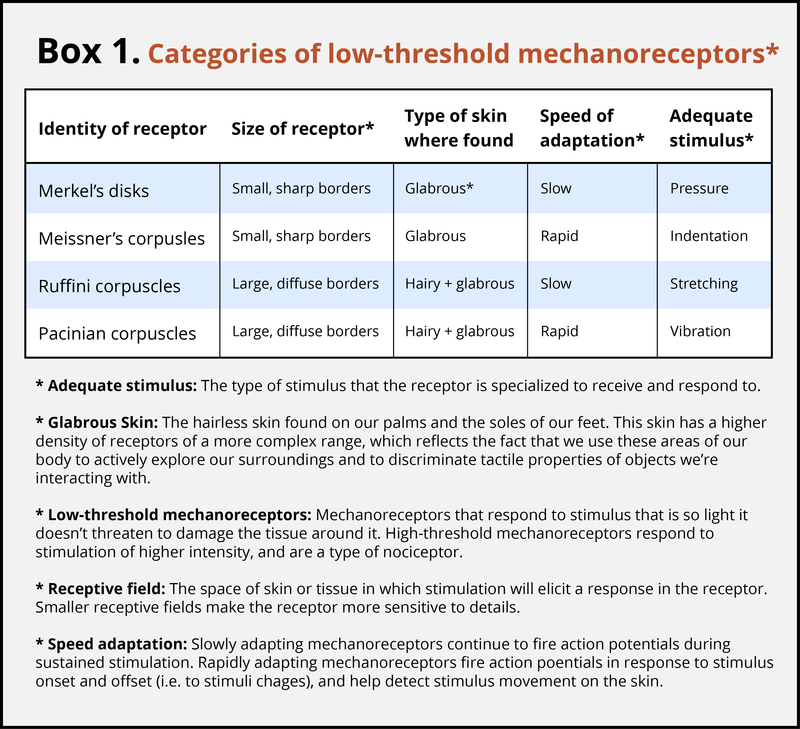
Action Potentials in the Receptor Cells Travel as Nerve Impulses with Different Speeds

Affective Aspects of Touch Are Important for Development and Relationships
Modulation
Pain Is Necessary for Survival, but Our Brain Can Stop It if It Needs To
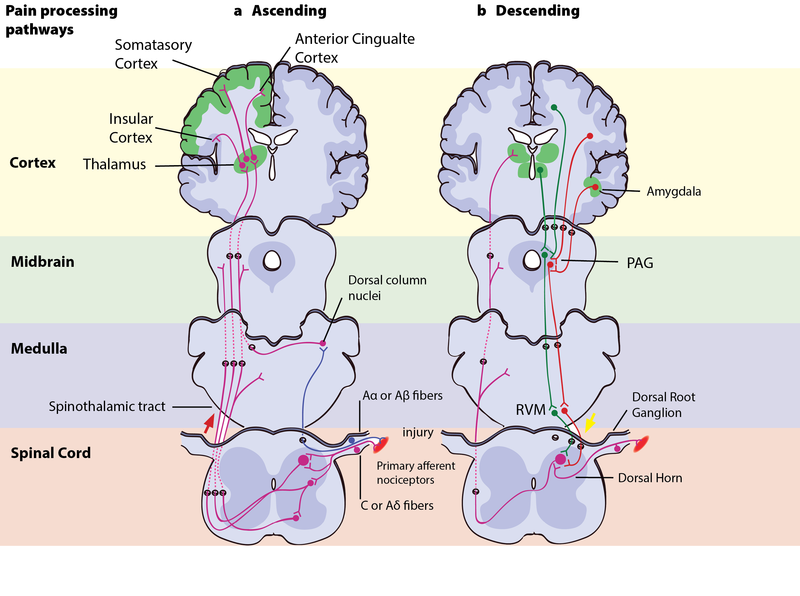
The Motivation–Decision Model and Descending Modulation of Pain
The Analgesic Power of Reward

When Touch Becomes Painful or Pain Becomes Chronic
The Power of the Mind

Summary
Outside Resources
- Book: Butler, D. S., Moseley, G. L., & Sunyata. (2003). Explain pain (p. 19). Australia: Noigroup.
- Book: Kringelbach, M. L., & Berridge, K. C. (Eds.). (2010). Pleasures of the brain (p. 343). Oxford, UK: Oxford University Press.
- Book: Ralston, A. (2004). Between a rock and a hard place: The basis of the motion picture 127 Hours. New York, NY: Atria.
- Book: Sacks, O. (1998). The man who mistook his wife for a hat: And other clinical tales. New York, NY: Simon & Schuster.
- Video: BBC Documentary series “Human Senses,” Episode 3: Touch and Vision
- watchdocumentary.org/watch/hu...f3e33c14a.html
- Video: BBC Documentary “Pleasure and Pain with Michael Mosley”
- http://www.bbc.co.uk/programmes/b00y377q
- Video: TEDxAdelaide - Lorimer Moseley – “Why Things Hurt”
- Video: Trailer for the film 127 Hours, directed by Danny Boyle and released in 2010
- Web: Homepage for the International Association for the Study of Pain
- http://www.iasp-pain.org
- Web: Proceedings of the National Academy of Sciences Colloquium "The Neurobiology of Pain"
- http://www.pnas.org/content/96/14.toc#COLLOQUIUM
- Web: Stanford School of Medicine Pain Management Center
- http://paincenter.stanford.edu/
- Website resource aiming to communicate “advances and issues in the clinical sciences as they relate to the role of the brain and mind in chronic pain disorders,” led by Dr. Lorimer Moseley
- www.bodyinmind.org/
Discussion Questions
Vocabulary
- A-fibers
- Fast-conducting sensory nerves with myelinated axons. Larger diameter and thicker myelin sheaths increases conduction speed. Aβ-fibers conduct touch signals from low-threshold mechanoreceptors with a velocity of 80 m/s and a diameter of 10 μm; Aδ-fibers have a diameter of 2.5 μm and conduct cold, noxious, and thermal signals at 12 m/s. The third and fastest conducting A-fiber is the Aα, which conducts proprioceptive information with a velocity of 120 m/s and a diameter of 20 μm.
- Allodynia
- Pain due to a stimulus that does not normally provoke pain, e.g., when a light, stroking touch feels painful.
- Analgesia
- Pain relief.
- C-fibers
- C-fibers: Slow-conducting unmyelinated thin sensory afferents with a diameter of 1 μm and a conduction velocity of approximately 1 m/s. C-pain fibers convey noxious, thermal, and heat signals; C-tactile fibers convey gentle touch, light stroking.
- Chronic pain
- Persistent or recurrent pain, beyond usual course of acute illness or injury; sometimes present without observable tissue damage or clear cause.
- C-pain or Aδ-fibers
- C-pain fibers convey noxious, thermal, and heat signals
- C-tactile fibers
- C-tactile fibers convey gentle touch, light stroking
- Cutaneous senses
- The senses of the skin: tactile, thermal, pruritic (itchy), painful, and pleasant.
- Descending pain modulatory system
- A top-down pain-modulating system able to inhibit or facilitate pain. The pathway produces analgesia by the release of endogenous opioids. Several brain structures and nuclei are part of this circuit, such as the frontal lobe areas of the anterior cingulate cortex, orbitofrontal cortex, and insular cortex; and nuclei in the amygdala and the hypothalamus, which all project to a structure in the midbrain called the periaqueductal grey (PAG). The PAG then controls ascending pain transmission from the afferent pain system indirectly through the rostral ventromedial medulla (RVM) in the brainstem, which uses ON- and OFF-cells to inhibit or facilitate nociceptive signals at the spinal dorsal horn.
- Endorphin
- An endogenous morphine-like peptide that binds to the opioid receptors in the brain and body; synthesized in the body’s nervous system.
- Exteroception
- The sense of the external world, of all stimulation originating from outside our own bodies.
- Interoception
- The sense of the physiological state of the body. Hunger, thirst, temperature, pain, and other sensations relevant to homeostasis. Visceral input such as heart rate, blood pressure, and digestive activity give rise to an experience of the body’s internal states and physiological reactions to external stimulation. This experience has been described as a representation of “the material me,” and it is hypothesized to be the foundation of subjective feelings, emotion, and self-awareness.
- Nociception
- The neural process of encoding noxious stimuli, the sensory input from nociceptors. Not necessarily painful, and crucially not necessary for the experience of pain.
- Nociceptors
- High-threshold sensory receptors of the peripheral somatosensory nervous system that are capable of transducing and encoding noxious stimuli. Nociceptors send information about actual or impending tissue damage to the brain. These signals can often lead to pain, but nociception and pain are not the same.
- Noxious stimulus
- A stimulus that is damaging or threatens damage to normal tissues.
- Ocial touch hypothesis
- Proposes that social touch is a distinct domain of touch. C-tactile afferents form a special pathway that distinguishes social touch from other types of touch by selectively firing in response to touch of social-affective relevance; thus sending affective information parallel to the discriminatory information from the Aβ-fibers. In this way, the socially relevant touch stands out from the rest as having special positive emotional value and is processed further in affect-related brain areas such as the insula.
- Pain
- Defined as “an unpleasant sensory and emotional experience associated with actual or potential tissue damage, or described in terms of such damage,” according to the International Association for the Study of Pain.
- Phantom pain
- Pain that appears to originate in an amputated limb.
- Placebo effect
- Effects from a treatment that are not caused by the physical properties of a treatment but by the meaning ascribed to it. These effects reflect the brain’s own activation of modulatory systems, which is triggered by positive expectation or desire for a successful treatment. Placebo analgesia is the most well-studied placebo effect and has been shown to depend, to a large degree, on opioid mechanisms. Placebo analgesia can be reversed by the pharmacological blocking of opioid receptors. The word “placebo” is probably derived from the Latin word “placebit” (“it will please”).
- Sensitization
- Increased responsiveness of nociceptive neurons to their normal input and/or recruitment of a response to normally subthreshold inputs. Clinically, sensitization may only be inferred indirectly from phenomena such as hyperalgesia or allodynia. Sensitization can occur in the central nervous system (central sensitization) or in the periphery (peripheral sensitization).
- S ocial touch hypothesis
- Proposes that social touch is a distinct domain of touch. C-tactile afferents form a special pathway that distinguishes social touch from other types of touch by selectively firing in response to touch of social-affective relevance; thus sending affective information parallel to the discriminatory information from the Aβ-fibers. In this way, the socially relevant touch stands out from the rest as having special positive emotional value and is processed further in affect-related brain areas such as the insula.
- Somatosensory cortex
- Consists of primary sensory cortex (S1) in the postcentral gyrus in the parietal lobes and secondary somatosensory cortex (S2), which is defined functionally and found in the upper bank of the lateral sulcus, called the parietal operculum. Somatosensory cortex also includes parts of the insular cortex.
- Somatotopically organized
- When the parts of the body that are represented in a particular brain region are organized topographically according to their physical location in the body (see Figure 8.5.2 illustration).
- Spinothalamic tract
- Runs through the spinal cord’s lateral column up to the thalamus. C-fibers enter the dorsal horn of the spinal cord and form a synapse with a neuron that then crosses over to the lateral column and becomes part of the spinothalamic tract.
- Transduction
- The mechanisms that convert stimuli into electrical signals that can be transmitted and processed by the nervous system. Physical or chemical stimulation creates action potentials in a receptor cell in the peripheral nervous system, which is then conducted along the axon to the central nervous system.