
9.1.2: Paleoenvironment and Hominin Evolution
- Last updated
- Save as PDF
- Page ID
- 136427
There is so much more to paleoanthropology than digging up and grouping fossil hominins: the discipline seeks to explain and understand the evolution of our ancestors’ behavior and morphology. There is no doubt that one of the major drivers (selective pressures) in hominin evolution is the environment. Large-scale changes in global and regional climate, as well as alterations to the environment, are all linked to hominin diversification, dispersal, and extinction (Maslin et al. 2014).
Environmental reconstructions often use modern analogues. Let us take, for instance, the hippopotamus. It is an animal that thrives in environments that have abundant water to keep its skin cool and moist. If the environment for some reason becomes drier, it is expected that hippopotamus populations will reduce. If a drier environment becomes wetter, it is possible that hippopotamus populations may be attracted to the new environment and thrive. Such instances have occurred multiple times in the past, and the bones of some fauna (animals, like the hippopotamus) that are sensitive to these changes give us insights into these events.
Reconstructing a paleoenvironment relies on a range of techniques, which vary depending on whether research interests focus on local changes or more global environmental changes/reconstructions. For local environments (reconstructing those of a single site or region), looking at the faunal assemblages (collections of fossils of other animals found at a site) and comparing them to animals found in certain modern environments allow us to determine if the environments in the past mirror those seen today in the region. Changes in the faunal assemblages, as well as when they occur and how they occur, tell us about past environmental changes. Other techniques are also useful in this regard. Isotopes of these fauna, for instance, tell us about the relative diets of individual fauna (e.g., using carbon isotopes to differentiate between species eating more grassland-heavy diets and those consuming bushland/tree-heavy diets) and whether the environment of individual animals was wetter or drier than the present day (e.g., nitrogen isotopes; Kingston & Harrison 2007).
Global climatic changes in the distant past, which fluctuated between being colder and drier and warmer and wetter on average, would have global implications for environmental change (Figure 9.3). These can be studied by using marine core and terrestrial soil data and by comparing these lines of evidence across multiple localities/sites/regions. These techniques allow us to use chemistry (such as nitrogen and oxygen isotopes in shells and sediments) or pollen grains (which show directly the kinds of flora surviving in an environment at a specific time period). This means that there are multiple lines of evidence that allow us to visualize global trends over millions of years (although it should be noted that the direction and extent of these changes could differ by geographic region).
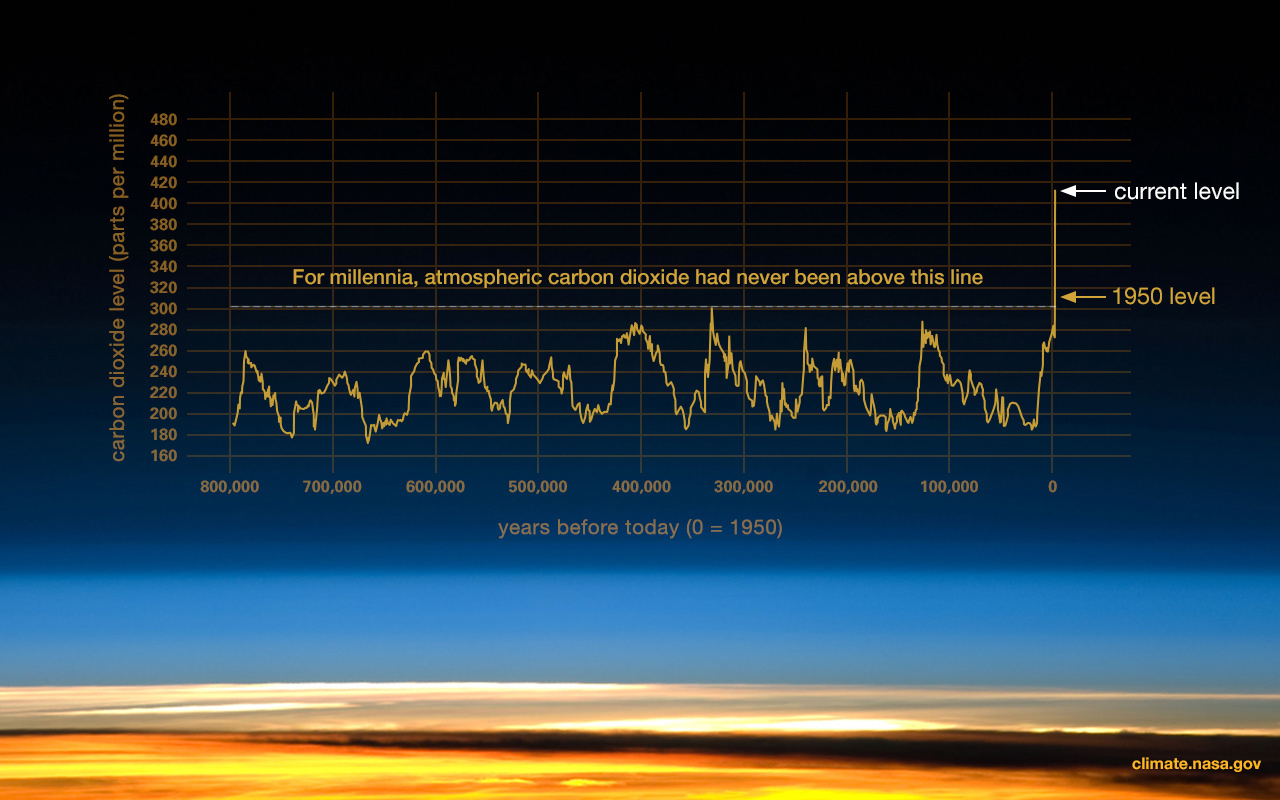 Figure \(\PageIndex{1}\): This graph, based on the comparison of atmospheric samples contained in ice cores and more recent direct measurements, illustrates how atmospheric CO₂ has fluctuated over time and increased sharply since the Industrial Revolution.
Figure \(\PageIndex{1}\): This graph, based on the comparison of atmospheric samples contained in ice cores and more recent direct measurements, illustrates how atmospheric CO₂ has fluctuated over time and increased sharply since the Industrial Revolution.Both local and global climatic/environmental changes have been used to understand parameters affecting our evolution (DeHeinzelin et al. 1999; Kingston 2007). There are numerous hypotheses regarding how climate has driven and continues to drive human evolution. Environmental change acts as an important keystone in hypotheses regarding the onset of several important hominin traits that are seen in early hominins and which are discussed in this chapter. Namely, the environment has been interpreted as:
- the driving force behind the evolution of bipedalism (terrestrial locomotion on two legs),
- the changing and diversifying of early hominin diets, and
- the diversification of multiple early hominin species.
Here, we will explore the five most popular hypotheses.
Savannah Hypothesis (or Aridity Hypothesis)
The hypothesis: This popular theory was first penned by Charles Darwin and supported by anthropologists like Raymond Dart (Darwin 1871; Dart 1925). It suggests that the expansion of the savannah (or less densely forested, drier environments) forced early hominins from an arboreal lifestyle (one living in trees) to a terrestrial one where bipedalism was a more efficient form of locomotion (Figure 9.4). This hypothesis stems from the idea that the Last Common Ancestor (LCA) between us and chimpanzees was a knuckle-walking quadruped like chimpanzees and gorillas. However, this idea was supported by little fossil or paleoenvironmental evidence and was later refined as the Aridity Hypothesis. The Aridity Hypothesis states that the long-term aridification and, thereby, expansion of savannah biomes were drivers in diversification in early hominin evolution (deMenocal and Bloemendal 1995; deMenocal 2004). It advocates particularly for periods of accelerated aridification leading to early hominin speciation events.
The evidence: While early bipedal hominins are often associated with wetter, more closed environments (i.e., not supporting the Savannah Hypothesis), both marine and terrestrial records seem to support general cooling, drying conditions, with isotopic records indicating an increase in grasslands (i.e., colder and wetter climatic conditions) between 8 mya and 6 mya across the African continent (Cerling et al. 2011). This can be contrasted with later climatic changes derived from aeolian dust records (sediments transported to the site of interest by wind), which demonstrate increases in seasonal rainfall between 3 mya and 2.6 mya, 1.8 mya and 1.6 mya, and 1.2 mya and 0.8 mya (deMenocal and Bloemendal 1995; deMenocal 2004).
Interpretation(s): Despite a relatively scarce early hominin record, it is clear that two important factors occur around the time period in which we see increasing aridity. The first factor is the diversification of taxa, where high morphological variation between specimens has led to the naming of multiple hominin genera and species. The second factor is the observation that the earliest hominin fossils appear to have traits associated with bipedalism and are dated to around the drying period (as based on isotopic records). Some have argued that it is more accurately a combination of bipedalism and arboreal locomotion, which will be discussed later. However, the local environments in which these early specimens are found (as based on the faunal assemblages) do not appear to have been dry.
 Figure \(\PageIndex{2}\): The African savannah grew during early hominin evolution. This may have forced early hominins from an arboreal lifestyle to a terrestrial one, where bipedalism was a more efficient form of locomotion.
Figure \(\PageIndex{2}\): The African savannah grew during early hominin evolution. This may have forced early hominins from an arboreal lifestyle to a terrestrial one, where bipedalism was a more efficient form of locomotion.Turnover Pulse Hypothesis
The hypothesis: In 1985, paleontologist Elisabeth Vbra noticed that in periods of extreme and rapid climate change, ungulates (hoofed mammals of various kinds) that had generalized diets fared better than those with specialized diets (Vrba 1988; Vrba 1998). Specialist eaters (eating and relying primarily on specific food types) faced extinction at greater rates than their generalist (able to eat more varied and variable diets) counterparts because they were unable to adapt to new environments. This meant that specialist eater species were often confined to isolated areas with dwindling resources, whereas generalist eaters were able to move out across the landscape in search of new food sources (Vrba 2000). Thus, periods with extreme climate change would be associated with high faunal turnover: that is, the extinction of many species and the speciation, diversification, and migration of many others to occupy various niches.
The evidence: The onset of the Quaternary Ice Age, between 2.5 mya and 3 mya, brought extreme global, cyclical interglacial and glacial periods (warmer, wetter periods with less ice at the poles, and colder, drier periods with more ice near the poles). Faunal evidence from the Turkana basin in East Africa indicates multiple instances of faunal turnover and extinction events, in which global climatic change resulted in changes from closed/forested to open/grassier habitats at single sites (Behrensmeyer et al. 1997; Bobe and Behrensmeyer 2004). Similarly, work in the Cape Floristic Belt of South Africa shows that extreme changes in climate play a role in extinction and migration in ungulates. While this theory was originally developed for ungulates, its proponents have argued that it can be applied to hominins as well. However, the link between climate and speciation is only vaguely understood (Faith and Behrensmeyer 2013).
Interpretation(s): While the evidence of rapid faunal turnover among ungulates during this time period appears clear, there is still some debate around its usefulness as applied to the paleoanthropological record. Specialist hominin species do appear to exist for long periods of time during this time period, yet it is also true that Homo, a generalist genus with a varied and adaptable diet, ultimately survives the majority of these fluctuations, and the specialists appear to go extinct.
Forest Hypothesis
The hypothesis: Based on contrasting environmental evidence to the Savannah hypothesis, R. J. Rayner and colleagues (1993) hypothesized that forested environments, rather than savannahs, were a key influence on the development of bipedalism in hominins. Unlike the Savannah Hypothesis, one criterion for this may be that the last common ancestor (LCA) between chimpanzees and us used an arboreal form of bipedal locomotion (i.e., walking along branches using the arms for stability), similar to orangutans, and was not a knuckle-walker like contemporary chimpanzees.
The evidence: Pollen evidence from the site of Makapansgat in South Africa indicated that around the time early hominins occupied the area, it was a closed, wooded environment. Similarly, the earliest evidence for bipedalism occurs in specimens (associated with taxa such as Orrorin and Ardipithecus spp. as well as Australopithecus anamensis) found in sites with evidence of closed habitats (Suwa et al. 2009). Furthermore, evidence of knuckle-walking in older hominin species is sorely lacking or highly contested.
Interpretation(s): This hypothesis can be considered in contrast to the Savannah Hypothesis, and it does appear to be evidence based. However, it is worth noting that preservation and resulting fossilization might be better in these kinds of environments, biasing this interpretation of the fossil record. Evidence for knuckle-walking in our more distant ancestors is also highly contested.
Variability Selection Hypothesis
The hypothesis: This hypothesis was first articulated by paleoanthropologist Richard Potts (1998). It links the high amount of climatic variability over the last 7 million years to both behavioral and morphological changes. Unlike previous notions, this hypothesis states that hominin evolution does not respond to habitat-specific changes or to specific aridity or moisture trends. Instead, long-term environmental unpredictability over time and space influenced morphological and behavioral adaptations that would help hominins survive, regardless of environmental context (Potts 1998; Potts 2013). The Variability Selection Hypothesis states that hominin groups would experience varying degrees of natural selection due to continually changing environments and potential group isolation. This would allow certain groups to develop genetic combinations that would increase their ability to survive in shifting environments. These populations would then have a genetic advantage over others that were forced into habitat-specific adaptations (Potts 2013).
The evidence: The evidence for this theory is similar to that for the Turnover Pulse Hypothesis: large climatic variability and higher survivability of generalists versus specialists. However, this hypothesis accommodates for larger time-scales of extinction and survival events.
Interpretation(s): In this way, the Variability Selection Hypothesis allows for a more flexible interpretation of the evolution of bipedalism in hominins, accommodating the discrepancies in evidence between the conflicting Savannah and Forest Hypotheses. This also allows for a more fluid interpretation of the Turnover Pulse Hypothesis, where species turnover is meant to be more rapid. In some ways, this hypothesis accommodates both environmental data and our interpretations of an evolution toward greater variability among species and the survivability of generalists.
Pulsed Variability Selection Hypothesis
The hypothesis: This hypothesis proposes that the East African Rift System (EARS) and changes in deep lakes are key drivers of diversification during early human evolution. EARS first developed about 10 mya and is responsible for the creation of large super lakes (e.g., Lake Baringo and Lake Turkana) within East Africa. The water contents of these lakes were primarily affected by both monsoonal rains and solar precessional cycles (cyclical changes in earth’s axis rotation- or wobble- that have global climatic effects). According to the Pulsed Variability Selection Hypothesis, human evolution was greatly affected by 200,000-year cyclical changes in aridity and humidity in this region, caused by those global cyclical changes .
The evidence: Proponents of this hypothesis name three extreme humid phases in East Africa at 2.7 mya to 2.5 mya, 1.8 mya to 1.6 mya, and 1 mya to 0.7 mya. During these periods, changes in solar precessional cycles increased the monsoonal system, causing more rain in East Africa, thereby increasing lake sizes. This is documented by the increase of diatomaceous lake sediments during these times. These expanded lakes would act as geographic barriers to hominin populations, allowing for changes and diversification in diet and adaptive behavior to the variable regions, even resulting in (allopatric) speciation (Maslin et al. 2014).
Interpretation(s): High levels of species diversity during these time periods as well as environmental indications of these barriers may allow for an interpretation of allopatric-speciation (i.e., speciation due to geographic barriers) events. However, the degree of interspecific variability and the extent to which these barriers acted as drivers of speciation are still debated.
Paleoenvironment Consolidated Summary
Some of the hypotheses presented in this section pay specific attention to habitat (Savannah and Forest Hypotheses) while others point to large-scale climatic forces (Pulsed Variability and Variability Selection Hypotheses). Some are complementary (Pulsed Variability and Turnover Pulse Hypotheses), whereas others are directly opposed to one another (Savannah and Forest Hypotheses). Some may be interpreted to describe the evolution of traits such as bipedalism (Savannah and Forest Hypotheses), and others more generally explain the diversification of early hominins (Turnover Pulse and Variability Selection Hypotheses). While there is no consensus as to how the environment drove our evolution, it is clear that the environment shaped both habitat and resource availability in ways that would have influenced our early ancestors physically and behaviorally.