
10.1.5: Homo Erectus- Biological and Cultural Innovations
- Last updated
- Save as PDF
- Page ID
- 136439
After 2 million years ago, a new hominin appeared on the scene. Known as Homo erectus, the prevailing scientific view was that this species was much more like us. These hominins were equipped with bigger brains and large bodies with limb proportions similar to our own. Perhaps most importantly, their way of life is now one that is recognizably human, with more advanced tools, hunting, use of fire, and colonizing new environments outside of Africa.
As will be apparent below, new data suggests that the story is not quite as simple. The fossil record for Homo erectus is much more abundant than that of Homo habilis, but it is also more complex and varied—both with regard to the fossils as well as the geographic context in which they are found. We will first summarize the anatomical characteristics that define Homo erectus, and then discuss the fossil evidence from Africa and the primary geographic regions outside Africa where the species has been located.
Homo erectus Anatomy
Compared to Homo habilis, Homo erectus showed increased brain size, smaller teeth, and a larger body. However, it also displayed key differences from later hominin species including our own.
Although the head of Homo erectus was less ape-like in appearance than the australopithecines, neither did it resemble modern humans (Figure 10.10). Compared to Homo habilis, Homo erectus had a larger brain size (average of about 900 cc compared to 650 cc to 750 cc). Instead of having a rounded shape like our skulls have, the erectus skull was long and low like a football, with a receding forehead, and a horizontal ridge called an occipital torus that gave the back of the skull a squared-off appearance. The cranial bones are thicker than those of modern humans, and some Homo erectus skulls have a slight thickening along the sagittal suture called a sagittal keel. Large, shelf-like brow ridges hang over the eyes. The face shows less prognathism, and the back teeth are smaller than those of Homo habilis. Instead of a pointed chin, like ours, the mandible of Homo erectus recedes back.
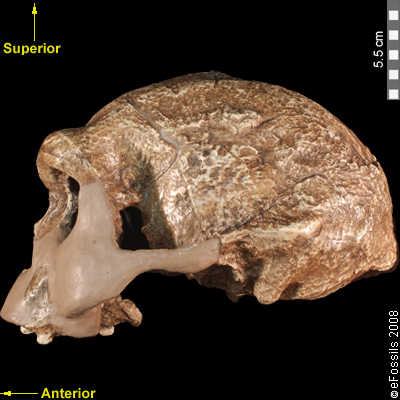 Figure \(\PageIndex{1}\): Replica of Homo erectus from Java, Indonesia. This cranium (known as Sangiran 17) dates to approximately 1.3 million to 1 million years ago. Note the large brow ridges and the occipital torus that gives the back of the skull a squared-off appearance.
Figure \(\PageIndex{1}\): Replica of Homo erectus from Java, Indonesia. This cranium (known as Sangiran 17) dates to approximately 1.3 million to 1 million years ago. Note the large brow ridges and the occipital torus that gives the back of the skull a squared-off appearance.Apart from these distinctive features, significant variation is present between Homo erectus fossils from different regions. Scientists have long noted differences between the fossils from Africa and those from Indonesia and China. For example, the Asian fossils tend to have a thicker skull and larger brow ridges than the African specimens, and the sagittal keel described above is more pronounced. Homo erectus fossils from the Republic of Georgia (described in the next section) also display distinctive characteristics. As with Homo habilis, this diversity has prompted a classification debate about whether or not Homo erectus should be split into multiple species. When African Homo erectus is characterized as a separate species, it is called Homo ergaster, while the Asian variant retains the erectus species name because it was discovered first. This text will use the species name Homo erectus for both variants.
Homo erectus was thought to have a body size and proportions more similar to modern humans. Unlike Homo habilis and the australopithecines, both of whom were small-statured with long arms and short legs, Homo erectus shows evidence of being fully committed to life on the ground. This meant long, powerfully muscled legs that enabled these hominins to cover more ground efficiently. Indeed, studies of the Homo erectus body form have linked several characteristics of the species to long-distance running in the more open savanna environment (Bramble and Lieberman 2004). Many experts think that hominins around this time had lost much of their body hair, were particularly efficient at sweating, and had darker-pigmented skin—all traits that would support the active lifestyle of such a large-bodied hominin (see Special Topic box).
Much of the information about the body form of Homo erectus comes from the Nariokotome fossil of the Homo erectus youth, described at the beginning of the chapter (see Figure 10.1). However, Homo erectus fossils are turning out to be more varied than previously thought. Homo erectus fossils from sites in Africa, as well as from Dmanisi, Georgia, show smaller body sizes than the Nariokotome boy’s. Even the Nariokotome skeleton itself has been reassessed to be quite a bit shorter (predicted to be closer to 5 feet 4 inches when fully grown, rather than over 6 feet), although there is still disagreement about which measurement is more accurate. One explanation for the range of body sizes could be adaptation to a range of different local environments, just as humans today show reduced body size in poor nutritional environments (Anton and Snodgrass 2012).
Homo erectus also shows some evidence of a reduction in sexual dimorphism in body size compared to the earlier australopithecines. In other words, Homo erectus males were only slightly larger in body size than females. The degree of sexual dimorphism among early hominin species is a contentious issue. It is a difficult characteristic to measure and assess in the fossil record, since fossils have to be complete enough to determine both body size and sex. However, if Homo erectus was less sexually dimorphic, it may signify changes in social organization within the species. If you recall from the chapter on primates, highly dimorphic species are those where males compete intensely for mating access to females. Decreased sexual dimorphism suggests that the lifestyle of Homo erectus may have been different from that of earlier hominins.
SPECIAL TOPICS: HOW WE BECAME HAIRLESS, SWEATY PRIMATES
As an anthropology instructor, one question about human evolution that students often ask me concerns human body hair—when did our ancestors lose it and why? It is assumed that our earliest ancestors were as hairy as modern-day apes. Today, though, we lack thick hair on most parts of our bodies except in the armpit and pubic regions and on the tops of our heads. Humans actually have about the same number of hair follicles per unit of skin as chimpanzees. But, the hairs on most of our body are so thin as to be practically invisible. When did we develop this peculiar pattern of hairlessness? Which selective pressures in our ancestral environment were responsible for this unusual characteristic?
Many experts believe that the driving force behind our loss of body hair was the need to effectively cool ourselves. Along with the lack of hair, humans are also distinguished by being exceptionally sweaty: we sweat larger quantities and more efficiently than any other primate. Humans have a larger amount of eccrine sweat glands than other primates and these glands generate an enormous volume of watery sweat. Sweating produces liquid on the skin that cools you off as it evaporates. It seems likely that hairlessness and sweating evolved together, as a recent DNA analysis has identified a shared genetic pathway between hair follicles and eccrine sweat gland production (Kamberov et al 2015).
Which particular environmental conditions led to such adaptations? In this chapter, we learned that the climate was a driving force behind many changes seen in the hominin lineage during the Pleistocene. At that time, the climate was increasingly arid and the forest canopy in parts of Africa was being replaced with a more open grassland environment, resulting in increased sun exposure for our ancestors. Compared to the earlier australopithecines, members of the genus Homo were also developing larger bodies and brains, starting to obtain meat by hunting or scavenging carcasses, and crafting sophisticated stone tools.
According to Nina Jablonski, an expert on the evolution of human skin, the loss of body hair and increased sweating capacity are part of the package of traits characterizing the genus Homo. While larger brains and long-legged bodies made it possible for humans to cover long distances while foraging, this new body form had to cool itself effectively to handle a more active lifestyle. Preventing the brain from overheating was especially critical. The ability to keep cool may have also enabled hominins to forage during the hottest part of the day, giving them an advantage over savanna predators, like lions, that typically rest during this time.
When did these changes occur? Although hair and soft tissue do not typically fossilize, there are several indirect methods that have been used to explore this question. One method tracks a human skin color gene. Since chimpanzees have light skin under their hair, it is probable that early hominins also had light skin color. Apes and other mammals with thick fur coats have protection against the sun’s rays. As our ancestors lost their fur, it is likely that increased melanin pigmentation was selected for to shield our ancestors from harmful ultraviolet radiation. A recent genetic analysis determined that one of the genes responsible for melanin production originated about 1.2 million years ago (Jablonski 2012).
Another line of evidence tracks the coevolution of a rather unpleasant human companion—the louse. A genetic study identified human body louse as the youngest of the three varieties of lice that infest humans, splitting off as a distinct variety around 70,000 years ago (Kittler, Kayser, and Stoneking 2003). Because human body lice can only spread through clothing, this may have been about the time when humans started to regularly wear clothing. However, the split between human head and pubic lice is estimated to have occurred much earlier, about three million years ago (Reed et al. 2007). When humans lost much of their body hair, lice that used to roam freely around the body were now confined to two areas: the head and pubic region. As a result of this “geographic” separation, the lice population split into two distinct groups.
Other explanations have also been suggested for the loss of human body hair. For example, being hairless has other advantages such as making it more difficult for skin parasites like lice, fleas, and ticks to live on us. Additionally, after bipedality evolved, hairless bodies would also make reproductive organs and female breasts more visible, suggesting that sexual selection may have played a role.
Homo erectus in Africa
Although the earliest discoveries of Homo erectus fossils were from Asia, the greatest quantity and best-preserved fossils of the species come from East African sites. The earliest fossils in Africa identified as Homo erectus come from the East African site of Koobi Fora, around Lake Turkana in Kenya, and are dated to about 1.8 million years ago. Other fossil remains have been found in East African sites in Kenya, Tanzania, and Ethiopia. Other notable African Homo erectus finds are a female pelvis from the site of Gona, Ethiopia (Simpson et al 2008), and a cranium from Olduvai Gorge known as Olduvai 9, thought to be about 1.4 million years old with massive brow ridges.
Homo erectus’ presence in South Africa is not well documented, though fossils thought to belong to the species have also been uncovered from the famed South African Swartkrans cave site along with stone tools and burned animal bones.
Regional Discoveries Outside Africa
It is generally agreed that Homo erectus was the first hominin to migrate out of Africa and colonize Asia and later Europe (although recent discoveries in Asia may challenge this view). Key locations and discoveries of Homo erectus fossils, along with the fossils’ estimated age are summarized below, and in Figure 10.12.
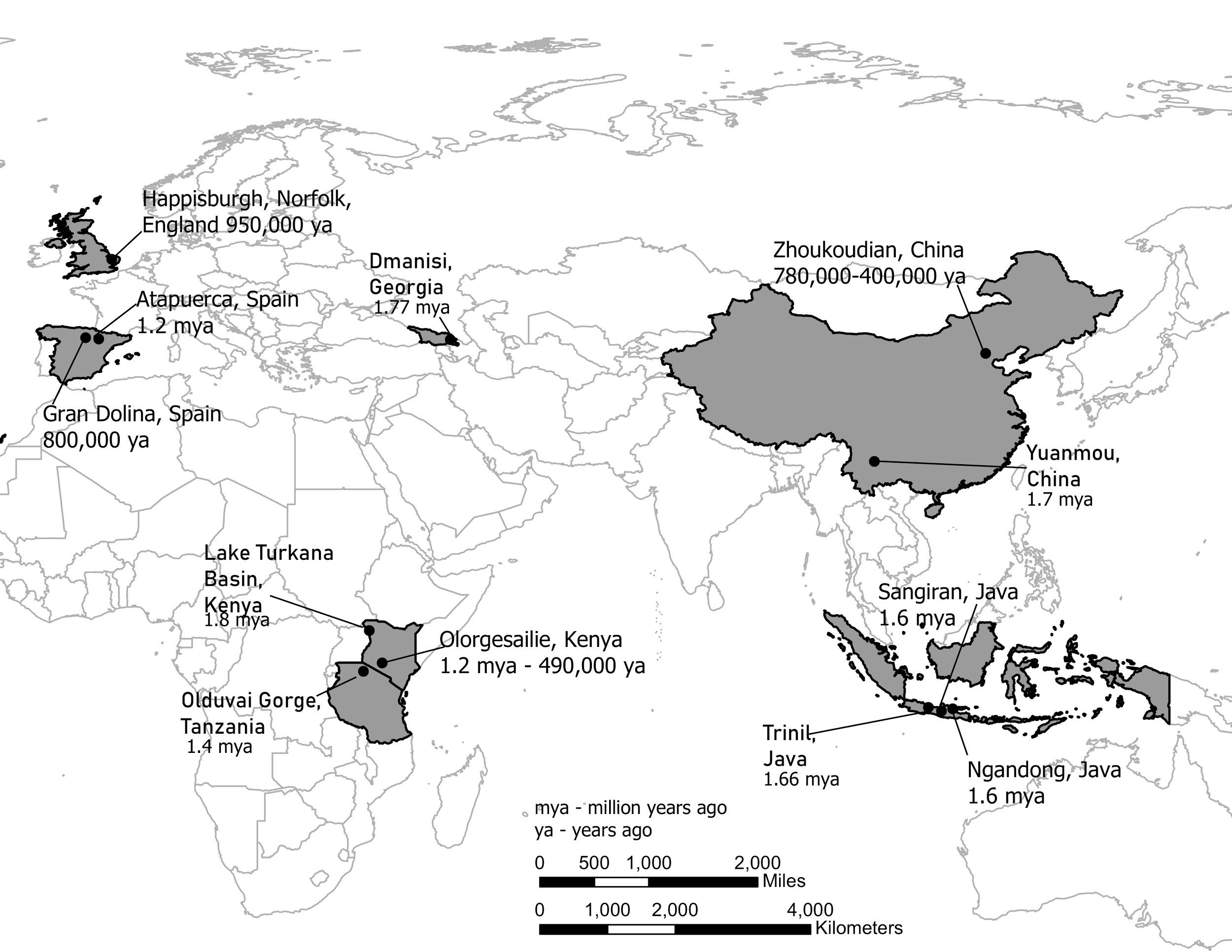 Figure \(\PageIndex{2}\): Map showing the locations of Homo erectus fossils around Africa and Eurasia.
Figure \(\PageIndex{2}\): Map showing the locations of Homo erectus fossils around Africa and Eurasia.Indonesia
The first discovery of Homo erectus was in the late 1800s in Java, Indonesia. A Dutch anatomist named Eugene Dubois searched for human fossils with the belief that since orangutans lived there, it might be a good place to look for remains of early humans. He discovered a portion of a skull, a femur, and some other bone fragments on a riverbank. While the femur looked human, the top of the skull was smaller and thicker than a modern person’s. Dubois named the fossil Pithecanthropus erectus (“upright ape-man”), popularized in the media at the time as “Java Man.” After later discoveries of similar fossils in China and Africa, they were combined into a single species (retaining the erectus name) under the genus Homo.
Homo erectus has a long history in Indonesia; further discoveries of fossils from Java were dated by argon dating to about 1.6 million to 1.8 million years. A cache of H. erectus fossils from the site of Ngandong in Java has yielded very recent dates of 43,000 years, although a more recent study with different dating methods concluded that they were much older—between 140,000 and 500,000 years old. Still, the possible existence of isolated, yet-to-be-discovered hominin populations in the region is of great interest to paleoanthropologists, especially given the discovery of the tiny Homo floresiensis fossils discovered on the nearby island of Flores, Indonesia, and the very recent announcement of possible tiny hominin fossils from the island of Luzon in the Philippines.
China
There is evidence of Homo erectus in China from several regions and time periods. Homo erectus fossils from northern China, collectively known as “Peking Man,” are some of the most famous human fossils in the world. Dated to about 400,000–700,000 years ago, they were excavated from the site of Zhoukoudian, near the outskirts of Beijing. Hundreds of bones and teeth, including six nearly complete skulls, were excavated from the cave in the 1920s and 1930s. Much of the fossils’ fame comes from the fact that they disappeared under mysterious circumstances. As Japan advanced into China during World War II, Chinese authorities, concerned for the security of the fossils, packed up the boxes and arranged for them to be transported to the United States. But in the chaos of the war, they vanished and were never heard about again. What exactly happened to them is murky—there are several conflicting accounts. Fortunately, an anatomist named Frans Weidenreich who had previously studied the bones had made casts and measurements of the skulls, so this valuable information was not lost. More recent excavations, at Longgushan “Dragon Bone Cave” at Zhoukoudian, of tools, living sites, and food remains, have revealed much about the lifestyle of Homo erectus during this time.
Despite this lengthy history of scientific research, China, compared to Africa, was perceived as somewhat peripheral to the study of hominin evolution. Although Homo erectus fossils have been found at several sites in China, with dates that make them comparable to those of Indonesian Homo erectus, none seemed to approximate the antiquity of African sites. The notable finds at sites like Nariokotome and Olorgesaille took center stage during the 1970s and 80’s, as scientists focused on elucidating the species’ anatomy and adaptations in its African homeland. In contrast, fewer research projects were focused on East Asian sites (Qiu 2016).
However, isolated claims of very ancient hominin occupation kept cropping up from different locations in Asia. While some were dismissed because of problems with dating methods or stratigraphic context, the 2018 publication of the discovery of stone tools from China dated to 2.1 million years caught everyone’s attention. Dated by paleomagnetic techniques that date the associated soils and windblown dust, these tools indicate that hominins in Asia predated those at Dmanisi by at least 300,000 years (Zhu et al. 2018). In fact, the tools are older than any Homo erectus fossils anywhere. Since no fossils were found with the tools, it isn’t known which species made them, but it opens up the intriguing possibility that hominins earlier than Homo erectus could have migrated out of Africa. These exciting new discoveries are shaking up previously held views of the East Asian human fossil record.
Western Eurasia
An extraordinary collection of fossils from the site of Dmanisi in the Republic of Georgia has revealed the presence of Homo erectus in Western Eurasia between 1.75 million and 1.86 million years ago. Dmanisi is located in the Caucasus mountains in Georgia. When archaeologists began excavating a medieval settlement near the town in the 1980s and came across the bones of extinct animals, they shifted their focus from the historic to the prehistoric era, but they probably did not anticipate going back quite so far in time! The first hominin fossils were discovered in the early 1990s, and since that time, at least five relatively well-preserved crania have been excavated.
There are several surprising things about the Dmanisi fossils. Compared to African Homo erectus, they have smaller brains and bodies. However, despite the small brain size, they show clear signs of Homo erectus traits such as heavy brow ridges and reduced facial prognathism. Paleoanthropologists have pointed to some aspects of their anatomy (such as the shoulders) that appear rather primitive, although their body proportions seem fully committed to terrestrial bipedalism. One explanation for these differences could be that the Dmanisi hominins represent a very early form of Homo erectus that left Africa before increases in brain and body size evolved in the African population.
Second, although the fossils at this location are from the same geological context, they show a great deal of variation in brain size and in facial features. One skull (Skull 5) has a cranial capacity of only 550 cc, smaller than many Homo habilis fossils, along with larger teeth and a protruding face. Scientists disagree on what these differences mean. Some contend that the Dmanisi fossils cannot all belong to a single species because each one is so different. Others assert that the variability of the Dmanisi fossils proves that they, along with all early Homo fossils, including H. habilis and H. rudolfensis, could all be grouped into Homo erectus (Lordikipanidze et al. 2013). Regardless of which point of view ends up dominating, the Dmanisi hominins are clearly central to the question of how to define the early members of the genus Homo.
Europe
Until recently, there was scant evidence of any Homo erectus presence in Europe, and it was assumed that hominins did not colonize Europe until much later than East Asia or Eurasia. One explanation for this was that the harsh ice age climate of Western Europe served as a barrier to living there. However, recent fossil finds from Spain suggest that Homo erectus could have made it into Europe over a million years ago. In 2008 a mandible from the Atapuerca region in Spain was discovered, dating to about 1.2 million years ago. A more extensive assemblage of fossils from the site of Gran Dolina in Atapuerca have been dated to about 800,000 years ago. In England in 2013 fossilized hominin footprints of adults and children dated to 950,000 years ago were found at the site of Happisburgh, Norfolk, which would make them the oldest human footprints found outside Africa (Ashton et al. 2014).
At this time, researchers aren’t in agreement as to whether the first Europeans belonged to Homo erectus proper or to a later descendent species. Some scientists refer to the early fossils from Spain by the species name, Homo antecessor.
|
Region |
Sites |
Dates |
Significance of Fossils |
|
East Africa |
East and West Lake Turkana, Kenya; Olduvai Gorge, Tanzania |
1.8 to 1.4 mya |
Earliest evidence of H. erectus; significant variation in skull and facial features. |
|
Western Eurasia |
Dmanisi, Republic of Georgia |
1.75 mya |
Smaller brains and bodies than H. erectus from other regions. |
|
Western Europe |
Atapuerca, Spain (Sima del Elefante and Gran Dolina caves) |
1.2 mya– 400,000 ya |
Partial jaw from Atapuerca is oldest evidence of H. erectus in Western Europe. Fossils from Gran Dolina (dated to about 800,000 years) sometimes referred to as H. antecessor. |
|
Indonesia |
Ngandong, Java; Sangiran, Java |
1.6 mya |
Early dispersal of H. erectus to East Asia; Asian H. erectus features. |
|
China |
Zhoukoudian, China; Loess Plateau (Lantian) |
780,000 – 400,000 ya 2.1 mya |
Large sample of H. erectus fossils and artifacts. Recent evidence of stone tools from Loess Plateau suggests great antiquity of Homo in East Asia. |