
14.1.2: Adjustments and Adaptations
- Last updated
- Save as PDF
- Page ID
- 136469
Adjustments
The term “adjustment” refers to an organism’s non-genetic way of coping with the stressors of its environment. Although adjustments themselves are non-genetic in nature, the ability of an organism to experience or develop an adjustment is based in its phenotypic plasticity, which is linked to its evolutionarily guided genetic potential. Adjustments occur exclusively on the individual level. As such, different individuals within a population may experience a wide range of possible adjustments in response to a similar stressor. In general, the three main forms of adjustment are: behavioral, acclimatory, and developmental.
Behavioral Adjustments
 Figure \(\PageIndex{1}\): Notice the lack of full spectrum color in this photo of a deep-water diver and the diver’s use of specialized equipment, such as a breathing apparatus to deliver gases for respiration, bodysuit to ensure thermal regulation, and flashlight to increase visibility in the low-light setting.
Figure \(\PageIndex{1}\): Notice the lack of full spectrum color in this photo of a deep-water diver and the diver’s use of specialized equipment, such as a breathing apparatus to deliver gases for respiration, bodysuit to ensure thermal regulation, and flashlight to increase visibility in the low-light setting.When you are cold, do you reach for a blanket? When you are warm, do you seek out shelter cooled by an air-conditioning system? If so, you have likely been influenced to do so by the culture in which you were raised. As noted earlier in the text, the term “culture” refers to a collection of shared, learned behaviors among individuals within a discrete population. Behavioral adjustments are regarded as cultural responses to environmental stressors. These adjustments are temporary in nature and, since they are nongenetic, must be constantly altered to meet novel situations posed by the environment. For example, through the use of a specialized mixture of gases for breathing, an apparatus for the delivery of the gases, protective clothing, and gear to increase visibility, divers are able to reach extraordinary depths (in excess of 300 meters below the surface) within the water. The deeper a diver descends, the more atmospheric pressure the diver experiences resulting in increased levels of potentially toxic byproducts of respiration within the body. In addition, with increased depth there is a decrease in the ambient temperature of the water as well as a decrease in the availability of light within the visible spectrum. Deep-water divers are well-versed in the environmental stressors of open waters and employ a variety of strategies based on behavioral adjustments to meet such demands. From wearing protective clothing to assist with maintaining the body’s core temperature to waiting at a specific depth for a prescribed period of time to facilitate the expulsion from the body of nitrogen gas that may have accumulated within the bloodstream, divers employ numerous behavioral adjustments to ensure their safety dive after dive (Figure 14.1). Without these culturally mediated behavioral adjustments, a deep-water diver’s first dive would be their last.
In many developing countries, the use of refrigeration for the storage of perishable food products is uncommon; therefore, individuals within these cultures have developed a variety of behavioral adjustment strategies related to food preparation to address the possibility of food spoilage. Through a cross-cultural analysis of spice use in recipes, Sherman and Billing (1999) determined that cultures closest to the equator, where temperatures are hotter, tend to use both a greater number and a wider variety of plant-based spices with bacteria-inhibiting phytochemical properties (e.g., garlic and onion) in their recipes than cultures located further from the equator. The antimicrobial properties of the spices permits the consumption of foods, particularly animal-based protein sources, for a period of time beyond that which would be considered safe. There are some acclimatory adjustment benefits to the use of some pungent spices as well, which are explored in the following section.
Acclimatory Adjustments: Thermal Stressors
Acclimatory adjustments are temporary, reversible changes in an organism’s physiology in response to environmental stressors. Although they are regarded as non-genetic in nature, the range of acclimatory adjustments an organism is capable of producing to accommodate a given stressor is linked to its underlying phenotypic plasticity and duration and severity of the stressor.
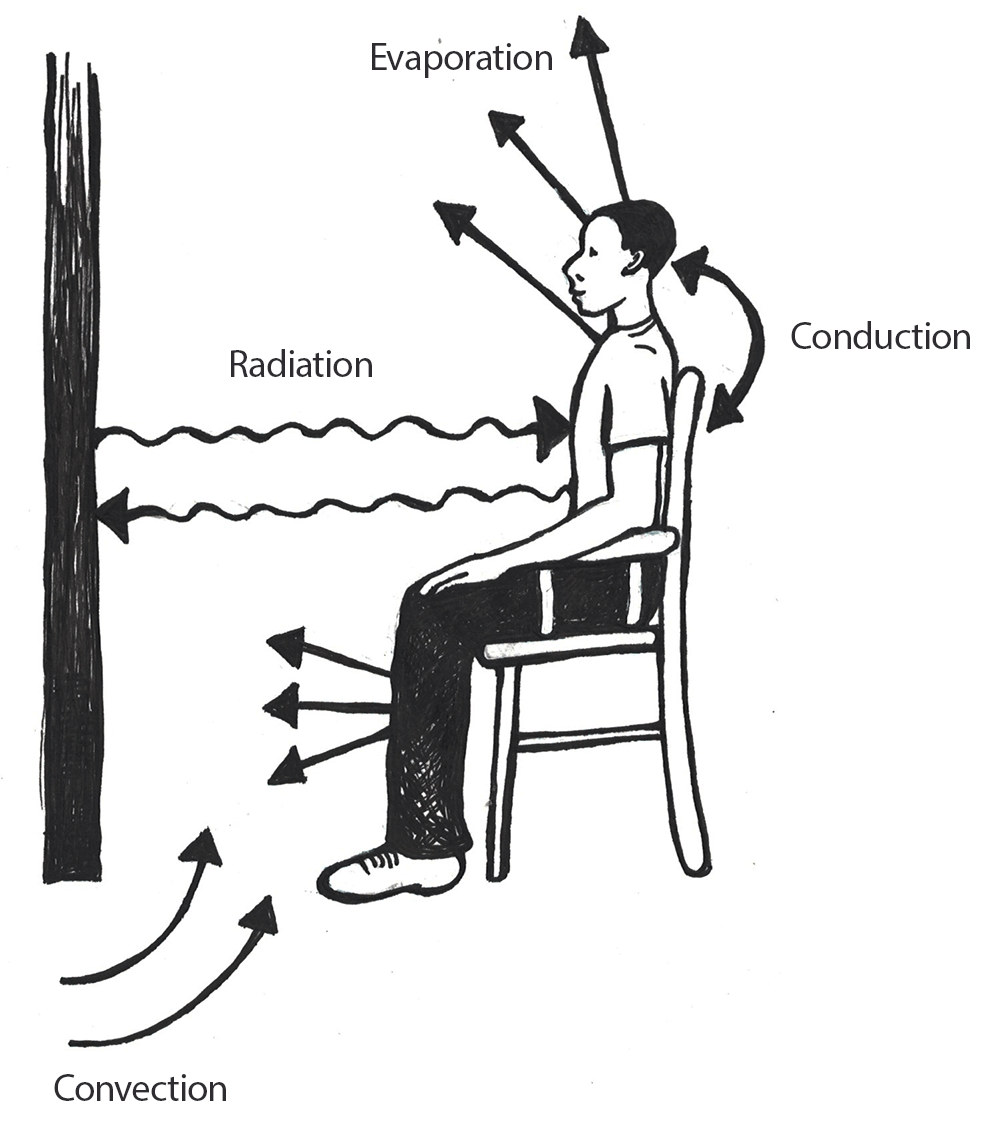 Figure \(\PageIndex{2}\): Various thermodynamic mechanisms related to heat gain and loss in the human body.
Figure \(\PageIndex{2}\): Various thermodynamic mechanisms related to heat gain and loss in the human body.Before we discuss how varying ambient temperatures affect the human body, we must detail the thermodynamic mechanisms through which heat may be gained or lost. There are four pathways for the loss of heat within the human body: conduction, convection, evaporation, and radiation (Figure 14.2).
Through conduction processes, heat will move from a warmer body to a cooler one through direct contact. An example of this is when you accidentally touch a hot cooktop with your hand and the heat is transferred from the cooktop to your skin.
With convection, when a warm body is surrounded by a cooler fluid (e.g. air or water) heat will be transferred from the warmer body to the cooler fluid. This is why we will often employ the behavioral adjustment of wearing multiple layers of clothing during the winter in an effort to prevent heat loss to the cooler atmosphere. Conversely, if your body temperature is cooler than that of the air surrounding you, your body will absorb heat.
Depending on your physical condition, most people will begin to sweat around 37.2℃ to 37.7℃ (98.9℉ – 99.9℉). Sweating is an example of evaporation, which occurs when a liquid, such as the water within our bodies, is converted to a gas. Phase conversions, such as those underlying the evaporative processes of transforming liquids to gases, require energy. In evaporation, this energy is in the form of heat, and the effect is to cool the body.
The final mechanism for heat loss within the human body is radiation, through which energy in the form of electromagnetic waves is produced at a wavelength that typically lies below that which is visible to the human eye. Although humans gain and lose heat from their bodies through radiation, this form of heat transfer is not visible. Humans are capable of losing and gaining heat through conduction, convection, and radiation; however, heat may not be gained through evaporation.
As the ambient temperature decreases, it becomes increasingly difficult for the human body to regulate its core temperature, which is central to the maintenance of homeostasis. The hypothalamus is a small portion of the human brain located near its base. It is responsible for numerous functions, including the regulation of body temperature. As measured orally, normal human body temperature averages 37℃ (98.6°F). When an individual’s body temperature falls below 34.4℃ (93.9°F), the hypothalamus becomes impaired leading to issues with body temperature control. A total loss of the ability to regulate body temperature occurs around 29.4℃ (84.9°F), which may result in death. When the ambient temperature (e.g. air temperature) falls below the critical temperature of 31℃ (87.8°F), a nude human body that is at rest will respond with a series of physiological changes to preserve homeostasis (Figure 14.3).
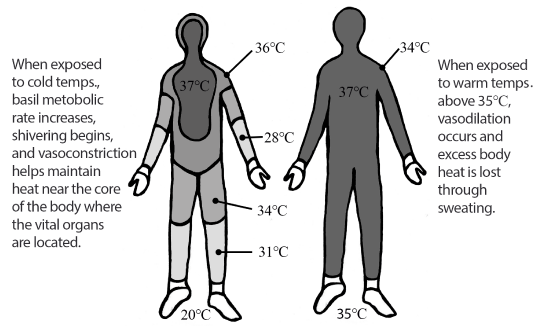 Figure \(\PageIndex{3}\): Example of overall body heat maintenance in cold and warm ambient environments.
Figure \(\PageIndex{3}\): Example of overall body heat maintenance in cold and warm ambient environments.In response to colder temperatures, the human body experiences two main types of physiological responses: those that increase the production of heat within the body and those that seek to retain the body’s heat. The production of heat within the body is accomplished through short-term increases in the body’s basal metabolic rate. This rate is a measure of the energy required to maintain necessary body processes while the body is in a resting state. As the body’s basal metabolic rate increases, an individual must consume greater quantities of energy-providing nutrients to maintain the increase. Of course, such increases may not continue forever as they are energetically expensive. As with all acclimatory adjustments, an increase in the basal metabolic rate is merely temporary.
Another form of a temporary heat-generating acclimatory adjustment to cold stress is the physiological response of shivering. Shivering results when the hypothalamus stimulates increased muscular activity that leads to an elevation of the muscular metabolism. Much like the increased muscular metabolism that occurs during periods of strenuous exercise, the elevation of muscular metabolism rates during shivering leads to higher rates of body heat generation.
Other physiological mechanisms the body uses to assist with the maintenance of temperature related to homeostasis involve the preservation of heat already contained within the body. Of these mechanisms, the most notable is the constriction of peripheral capillaries in the skin through a process called vasoconstriction. The decreased surface area of the capillaries through vasoconstriction results in less heat reaching the surface of the skin where it would be dissipated into the atmosphere. In addition, vasoconstriction leads to the maintenance of heat near the core of the body where the vital organs are located. This is one of the reasons that an individual may experience cold-related injuries, such as frost-bite leading to tissue necrosis (tissue death) in regions of the body that are most distant from the core (e.g. fingers, toes, nose, ears, cheeks, chin, etc.).
Just as cold stress presents challenges to maintaining homeostasis within the body with respect to temperature, heat does as well. In hot climates, the body will begin to absorb extra heat from its surroundings (through conduction, convection, and radiation) resulting in potential heat-related disorders, such as heat exhaustion. When the human body is exposed to ambient temperatures above 35℃ (95°F), excess body heat will be lost primarily through evaporative processes, specifically through sweating. All humans, regardless of their environment, have approximately the same number of sweat glands within their bodies. Over time, individuals living in hot, arid environments will develop more sensitive forms of sweat glands resulting in the production of greater quantities of sweat. In an effort to prevent dehydration due to this form of acclimatory adjustment, there will be an additional reduction in the volume of urine produced by the individual.
As noted in the previous section, some cultural groups, particularly those in equatorial regions, add pungent spices to their foods to inhibit the colonization of bacteria (Sherman and Billing 1999). Although the addition of spices to foods to decrease spoilage rates is a behavioral adjustment, the application of some forms of peppers triggers an acclimatory adjustment process as well. Compounds referred to as capsaicinoids are the secondary byproducts of chili pepper plants’ metabolism. Capsaicinoids are produced by the plants to deter their consumption by some forms of fungi and mammals. When mammals, such as humans, consume the capsaicinoids from chili peppers, a burning sensation may occur within their mouths and along their digestive tracts. This burning sensation is the result of the activation of capsaicin receptors along the body’s nerve pathways. Although the peppers themselves may be at ambient temperature so their consumption is technically not causing any form of body temperature increase, the human body perceives the pepper as elevating its core temperature due to the activation of the capsaicin receptors. Even though the body’s temperature has not actually been elevated due to the consumption of capsaicin, the hypothalamus will react as if it has, which leads to the initiation of sweating processes in an attempt to lower body temperature and maintain homeostasis. The increased piquancy (application of pungent spices to food) as a means of inhibiting food-borne bacterial colonization in warm climates, as well as spices’ ability to trigger sweating processes as a method for cooling the body, is an example of the intersection between behavioral and acclimatory adjustments that may be utilized by individuals within certain populations.
In addition to increased sweat production in the body as a means of regulating internal body temperature to maintain homeostasis, vasodilation may occur (Figure 14.4). Vasodilation occurs when there is an expansion of the capillaries within the skin leading to a more effective transfer of heat from within the body to the exterior to allow conductive, convective, radiative, and evaporative (sweating) processes to occur.
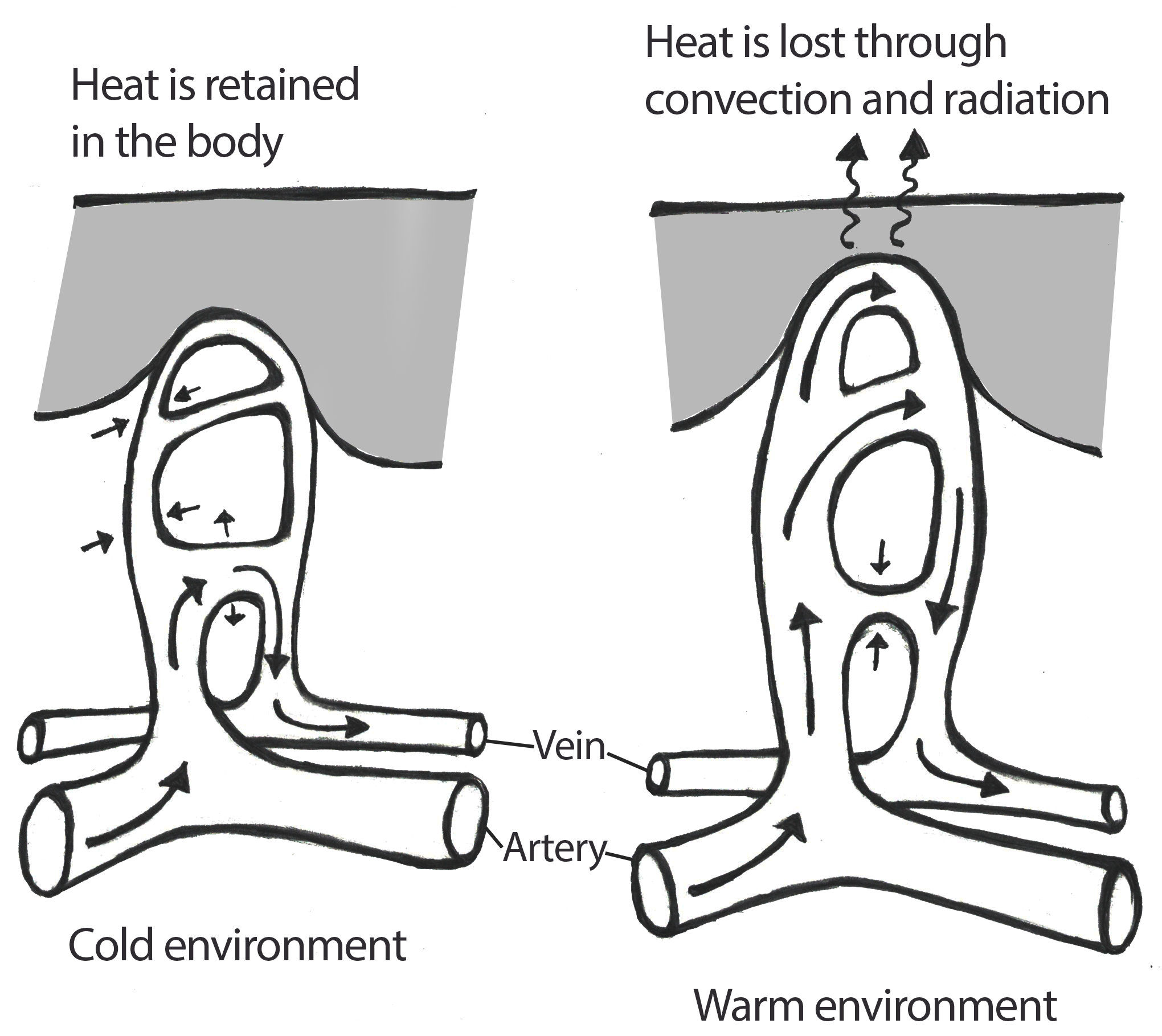 Figure \(\PageIndex{4}\): This image features an illustration of the vasoconstriction processes that occur within the peripheral vascular system when an individual is exposed to cold ambient temperatures and the vasodilation that occurs in warmer environments.
Figure \(\PageIndex{4}\): This image features an illustration of the vasoconstriction processes that occur within the peripheral vascular system when an individual is exposed to cold ambient temperatures and the vasodilation that occurs in warmer environments.Physiologically-based acclimatory adjustments to hot, dry climates may be complemented by behavioral adjustments as well. For example, individuals in such climates may limit their physical activity during the times of day when the temperature is typically the hottest. Additionally, these individuals may wear loose fitting clothing that covers much of their skin. The looseness of the clothing allows for air to flow between the clothing and the skin to permit the effective evaporation of sweat. Although it may seem counterintuitive to cover one’s body completely in a hot climate, the covering of the skin keeps the sun’s rays from directly penetrating the skin and elevating the body’s core temperature.
Acclimatory Adjustments: Altitudinal Stressors
The challenges posed by thermal conditions are but one form of environmental stressor humans must face. High-altitude environments, which are defined as altitudes in excess of 2400 meters above sea level (m.a.s.l.) or 7874 feet above sea level (f.a.s.l.), pose additional challenges to the maintenance of homeostasis in humans. Some of the main stressors encountered by those living within high-altitude environments include: decreased oxygen availability, cold temperatures, low humidity, high wind speed, a reduced nutritional base, and increased solar radiation levels. Of these challenges, the most significant is the decreased availability of oxygen.
 Figure \(\PageIndex{5}\): As altitude increases, atmospheric pressure decreases, which allows for more space between air molecules.
Figure \(\PageIndex{5}\): As altitude increases, atmospheric pressure decreases, which allows for more space between air molecules.To visualize how altitude affects the availability of oxygen, imagine two balloons that are each filled with the same quantity of oxygen molecules. One of these balloons is positioned at sea-level and the other is placed high upon a mountain peak. For the balloon at sea level, there is more atmospheric pressure pressing down on the molecules within this balloon. This leads to the oxygen molecules within the sea level balloon being forced into a more compact organization. In contrast, the mountain peak balloon has less atmospheric pressure pressing down on it. This leads to the oxygen molecules within that balloon spreading out from each other since they are not being forced together quite as strongly. This example highlights the availability of oxygen molecules in each breath than we take in low- versus high-altitude environments. At 5,500 m.a.s.l. (approximately 18,000 f.a.s.l.), the atmospheric pressure is approximately 50% of its value at sea level (Peacock 1998). At the peak of Mount Everest (8,900 m.a.s.l. or approximately 29,200 f.a.s.l.), the atmospheric pressure is equivalent to only about 30% of their sea level amounts (Peacock 1998) (Figure 14.5).
Due to the decreased availability of oxygen at higher altitudes, certain acclimatory adjustments are required to ensure the maintenance of homeostasis for individuals other than those who were gestated, born, and raised at high altitude. For these people, their rate of breathing will increase to permit greater quantities of air containing oxygen into the lungs when they ascend into higher altitude environments. An increased speed and depth of breathing, which is referred to as hyperpnea, is not sustainable indefinitely; thus, the rate of breathing begins to decrease as the person becomes acclimatized to the altitude. During the initial phases of high-altitude-related hyperpnea, the heart will begin to beat faster but the stroke volume (the amount of blood pushed through during each beat) will decrease slightly. In addition, the body will divert energy from non-critical bodily functions, such as digestive processes.
Once the atmospheric oxygen reaches the alveoli (small air sacs) in the lungs, it diffuses (spreads) across the alveolar membrane and enters erythrocytes (red blood cells). When the oxygen reaches the erythrocytes, it will loosely bind with hemoglobin, which is an iron-rich protein. It is within the alveoli that the oxygen combines with the hemoglobin. When the erythrocytes carrying the hemoglobin-binded oxygen molecules reach the capillaries where the partial pressure of oxygen is relatively low, the oxygen will be released by the hemoglobin so that it is free for diffusion into body cells. High-altitude-related hyperpnea leads to an increase in the pH of the blood, which makes the blood more alkaline. An increase in the alkalinity of blood is directly related to the partial pressures of oxygen and carbon dioxide in the bloodstream (Shah et al. 2006).
Similar to the acclimatory adjustments related to thermal conditions (e.g., shivering or sweating), those related to high altitude may not be sustained permanently due to their energetically expensive nature. Over a period of days or weeks, the human body will begin to compensate for the increases in respiration as well as pH through the urinary excretion of bicarbonate (a metabolic byproduct). In addition, to assist with the transportation of oxygen in the absence of decreased respiration, there will be an increase in the following: hematocrit (percentage volume of erythrocytes in the blood), myoglobin (oxygen- and iron-binding protein in muscle tissues), red blood cell mass, pulmonary artery pressure, and quantity of capillaries in the skeletal muscular tissue.
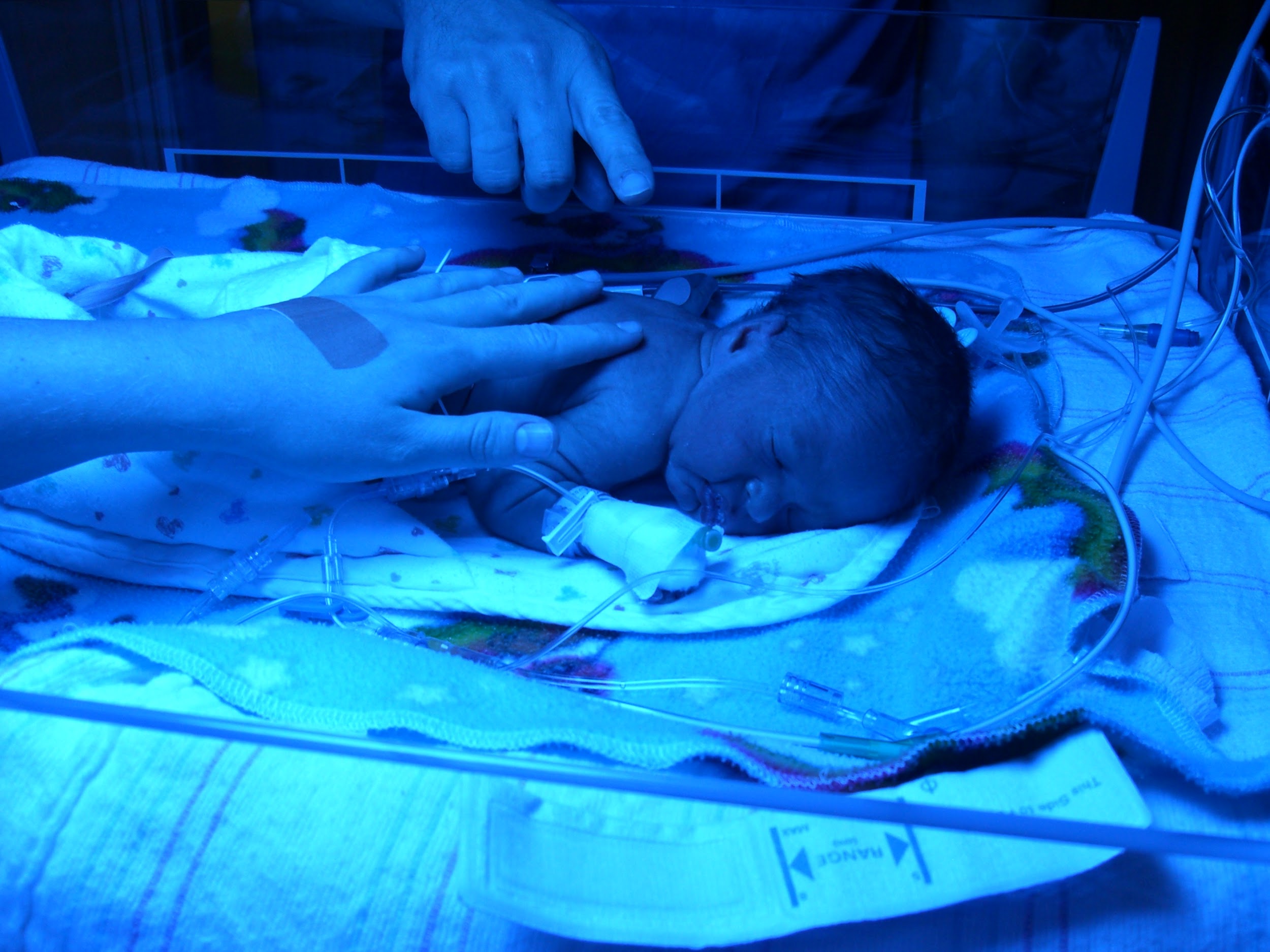 Figure \(\PageIndex{6}\): Premature infant born at 30 weeks, 4 days gestation to a mother with altitudinal-induced preeclampsia. Blue light assists the infant’s liver with processing high levels of bilirubin.
Figure \(\PageIndex{6}\): Premature infant born at 30 weeks, 4 days gestation to a mother with altitudinal-induced preeclampsia. Blue light assists the infant’s liver with processing high levels of bilirubin.Although the long-term acclimatory adjustments that an individual from low altitude experiences in a high-altitude environment may permit them to reside there successfully, reproduction within such settings is frequently complicated. With increased altitude comes an increased risk of miscarriage, lower birth weights, and higher infant mortality rates. As the mother’s body seeks to preserve its own homeostasis when faced with challenges related to high-altitude living, there is often a decreased rate and volume of blood flow to the uterus as compared to a pregnant woman of similar physiological condition at a lower altitude (Moore et al. 1998). Of course, a decreased rate and volume of uterine blood flow results in a decrease in the amount of oxygen that will be passed through the uterus and placenta to the developing fetus. In addition, women who experience pregnancy at higher altitudes are more prone to developing preeclampsia (severe elevation of blood pressure), which is linked to increased rates of both fetal and maternal death (Moore et al. 1998) (Figure 14.6).
Developmental Adjustments
Although there are often significant challenges for a mother gestating a fetus at high altitude, individuals who are native to such environments experience a form of adjustment referred to as a developmental adjustment. Developmental adjustments occur only in individuals who spent their developmental period (i.e. childhood and adolescence) within a high-altitude environment and they do not apply to those who moved into these environments in the post-developmental (i.e., adult) phase. Furthermore, the degree of developmental adjustment within an individual is directly related to their underlying phenotypic plasticity (polyphenism) as well as the amount of time during the crucial growth and development period the individual resides within the challenging environment. Although humans have the remarkable capacity to develop and survive within environments that are not overly conducive to the successful maintenance of homeostasis, there are definitely physiological costs associated with this ability.
 Figure \(\PageIndex{7}\): Quechua woman from high-altitude region of the Peruvian Andes.
Figure \(\PageIndex{7}\): Quechua woman from high-altitude region of the Peruvian Andes.In general, high-altitude natives tend to grow more slowly and physically mature later than their low-altitude counterparts (Figure 14.7). Decreased growth and maturity rates are linked not only to the increased physiological demands placed on the body due to the decreased partial pressure of oxygen but to decreases in the quality of the nutritional base at higher altitudes. Increased terrain complexity, elevated solar radiation levels, and higher wind speeds coupled with decreased temperatures and lower humidity levels at higher altitudes leads to difficulties with growing and maintaining crops and raising livestock. Overall, there is a decreased quality in the available nutritional base as altitude increases, which is correlated to a lack of the nutrients necessary to ensure proper physiological growth and development in humans. Thus, even though individuals may be able to develop and grow within high-altitude environments, they may not reach their full genetically mediated growth potential as they would in a lower-altitude environment.
The heart and lung capacity of individuals who are lifelong residents of high-altitude environments is larger than for individuals from lower-altitude regions. In addition, the high-altitude individuals are more efficient than those from low altitudes at diffusing oxygen from the bloodstream to the body’s various tissues. Of course, as noted previously, the developmental adjustments that an individual may experience are defined by their underlying genetic composition (phenotypic plasticity) and researchers have begun to discover some of the genetic factors that appear to be unique to certain populations of humans who have resided in high-altitude environments for significant periods of time. The time frame necessary for natural selection processes to act upon the underlying genetic composition of a population is on the order of many millennia; thus, the physiological changes to indigenous high-altitude populations on a whole are best described in the following section on adaptations.
Not all developmental adjustments are linked to environmental pressures such as climate or altitude; rather, some of these adjustments are correlated to sociocultural or behavioral practices. As noted earlier, some behavioral adjustments permit individuals to maintain homeostasis in challenging environments (e.g., wearing heavy clothing layers to maintain body temperature in cold weather). Other behavioral adjustments, many of which are based on sociocultural principles, may affect the physiological appearance of an individual when they are practiced consistently during the development and growth phases.
Sudden infant death syndrome (SIDS) has no definitive cause; however, the American Academy of Pediatrics published a report in 1992 linking SIDS to infants (under the age of one) sleeping on their stomachs. The “Back to Sleep” campaign championed by the American Academy of Pediatrics helped educate members of the medical community as well as the public that the best sleep position for infants is on their backs. Per the American Academy of Pediatrics Task Force on Infant Sleep Position and Sudden Infant Death Syndrome (2000: 1,245), between 1992 and 2000 the frequency of infants being placed on their stomachs to sleep decreased from more than 70% to less than 20% and the SIDS rate decreased by more than 40%.
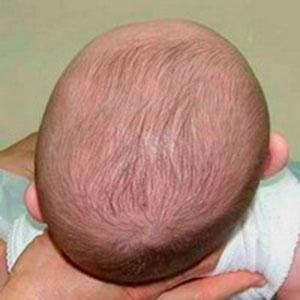 Figure \(\PageIndex{8}\): Infant with positional plagiocephaly. Notice the irregular shape of the posterior (back) of the skull.
Figure \(\PageIndex{8}\): Infant with positional plagiocephaly. Notice the irregular shape of the posterior (back) of the skull.Placing infants on their backs to sleep has led to decreased infant mortality (death) rates due to SIDS; however, it has led to an unintended consequence: infant cranial deformation. The cranial deformations experienced by infants who sleep solely on their back tend to manifest in one of two forms: brachycephaly and plagiocephaly (Roby et al. 2012). With positional brachycephaly, the back of the infant’s head appears rather uniformly flattened due to repetitive contact with a flat surface, such as a crib mattress or car seat back. In cases of positional plagiocephaly, the back of the infant’s head appears asymmetrically flattened (Figure 14.8). This asymmetry is typically due to an uneven distribution of mechanical forces resulting from the manner in which the infant’s head is in contact with a flat surface. The forms of cranial deformation resulting from sleep positioning do not affect the infant’s brain development. For many individuals, the appearance of the deformation is minimized during later development. Still, some individuals will maintain the pattern of cranial deformation acquired during their infancy throughout their lives. The unintentional cranial deformation resulting from placing infants on their backs to sleep as a means of preventing SIDS-related deaths is a physiological indicator of a behavioral adjustment.
Adaptations
As we have just explored, survival and reproduction at high altitudes present numerous physiological challenges for most humans, but what if there were some humans who were specially adapted to life at high altitudes? The behavioral, acclimatory, and developmental adjustments discussed above are all related to the phenotypic plasticity of the individual; however, most adjustments are temporary in nature and they affect a single individual rather than all individuals within a population. But, what if the physiological changes were permanent? What if they affected all members of a population rather than just a single individual? The long-term, micro-evolutionary (i.e., genetic) changes that occur within a population in response to an environmental stressor are referred to as an adaptation. From an evolutionary standpoint, the term “adaptation” refers to a phenotypic trait (i.e., physiological/morphological feature or behavior) that has been acted upon by natural selection processes to increase a species’ ability to survive and reproduce within a specific environment. Within the field of physiology, the term “adaptation” refers to traits that serve to restore homeostasis. The physiology-based interpretation of adaptations presumes that all traits serve a purpose and that all adaptations are beneficial in nature; however, this may be a fallacy, since some traits may be present without clear evidence as to their purpose. As such, during the following discussion of various forms of adaptations in human populations, we will focus our attention to phenotypic traits with an evidence-based purpose.
Adaptation: Altitudinal Adaptation
As mentioned in the previous section, there is genomic research supporting the evolutionary selection of certain phenotypes and their corresponding genotypes within indigenous high-altitude populations across the globe. The following discussion focuses on three high-altitude indigenous populations from Tibet, the Andes, and Ethiopia (Figure 14.9). Although these populations share many common genetic traits based on relatively similar evolutionary histories influenced by similar environmental stressors, there is support for local genetically based adaptation as well, based on different genes being acted upon by environmental stressors that may be unique to Tibet, the Andes, and Ethiopia (Bigham 2016).
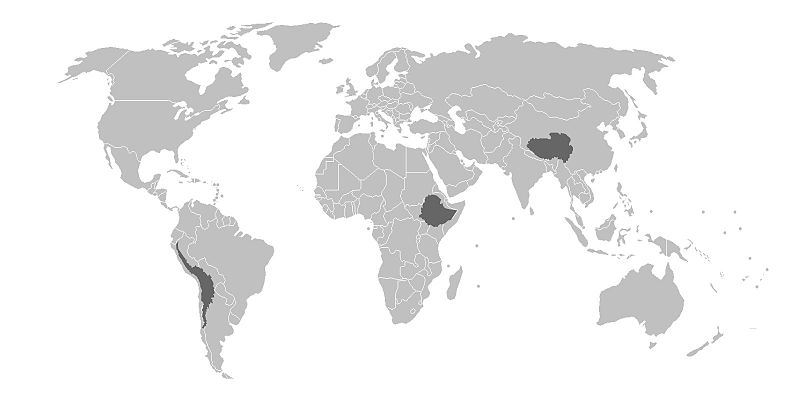 Figure \(\PageIndex{10}\): Highlighted regions feature (from left to right) the Andean, Simian (Ethiopian), and Tibetan Plateau high-altitude regions.
Figure \(\PageIndex{10}\): Highlighted regions feature (from left to right) the Andean, Simian (Ethiopian), and Tibetan Plateau high-altitude regions.Tibetan populations have resided in the Tibetan Plateau and Himalayan Mountain regions at elevations exceeding 4,000 m.a.s.l. (13,100 f.a.s.l.) for at least the past 7,400 years (Meyer et al. 2017). A gene referred to as EPAS1 is involved in the regulation of red blood cell (and hemoglobin) production as well as catecholamine homeostasis. Catecholamines are hormones (e.g., epinephrine) secreted as part of the sympathetic nervous system’s acute stress (“fight-or-flight”) response. An acute stress response typically includes an increase in: heart rate, blood pressure, and blood glucose levels. Long-term elevation of catecholamines in the body may lead to hypertension (elevated blood pressure), increased blood pH levels, the development of a form of cardiovascular disease leading to narrowing of the arteries (atherosclerosis), and blood clots. In the short term, an acclimatory adjustment leading to the increase of catecholamines and hemoglobin production by the hypoxia (low-oxygen level)-induced activation of the EPAS1 gene may assist individuals from lower altitudes as they ascend to significantly higher altitudes; however, such increases may not be maintained for long before they cause damage to the body. For indigenous high-altitude populations of Tibet, a mutation in the EPAS1 gene inhibits increased red blood cell production and assists with catecholamine regulation. The red blood cell count of high-altitude Tibetans with the EPAS1 point mutation is about the same as for individuals residing at sea level.
Interestingly, individuals in populations from the high-altitude Andean Altiplano of Peru and Bolivia, such as the Quechua and Aymara, lack the EPAS1 point mutation, so their red blood cell counts are relatively elevated compared with the Tibetan populations. In addition, populations from the Andean Altiplano also have an increased arterial oxygen saturation and low, hypoxia-induced breathing pattern (minor hyperventilation) as compared to their Tibetan counterparts (Bigham 2016). Populations indigenous to the Semien Plateau of Ethiopia, such as the Oromo and Amhara, share a similar but not identical EPAS1 point mutation with the Tibetan population (Bigham 2016). This suggests that the EPAS1 mutations occurred independently from each other; however, their effects are still similar in that they permit the Tibetan and Ethiopian populations to survive at high altitudes. Not all adaptations are related to life in high-altitude environments, however. In the following sections, we will address two more general examples of adaptation in human populations: variations in skin color and differences in body build.
Adaptation: Skin Tone Basics
When you think about your own skin tone and compare it to members of your family, do you all possess exactly the same shade? Are some members of your family darker than others? What about your friends? Your classmates? Skin tone occurs along a continuum, which is a reflection of the complex evolutionary history of our species. The expression of skin tone is regulated primarily by both melanin and hemoglobin. Melanin is a dark brown-black pigment that is produced by the oxidation of certain amino acids (e.g., tyrosine, cysteine, phenylalanine) in the melanocytes. Melanocytes are specialized cells located in the base layer (stratum basale) of the skin’s epidermis as well as several other areas within the body (Figure 14.10). Within the melanocytes, melanin is produced in the special organelle called a melanosome. Melanosomes serve as sites for the synthesis, storage, and transportation of melanin. Melanosomes transport the melanin particles through cellular projections to epidermal skin cells (keratinocytes) as well as to the base of the growing hair root (root sheath portion). In the eye, however, melanin particles produced by the melanosomes remain present within the iris (iridial melanocytes) and are not transported beyond their origin location. The two main forms of melanin related to skin, hair, and eye color are eumelanin and pheomelanin. All humans contain both eumelanin and pheomelanin within their bodies; however, the relative expression of these two forms of melanin determines an individual’s overall coloring. Eumelanin is a brown-to-black colored melanin particle and pheomelanin is more pink-to-red. Individuals with darker skin or hair color have a greater expression of eumelanin than those with lighter-colored skin and blonde or red hair.
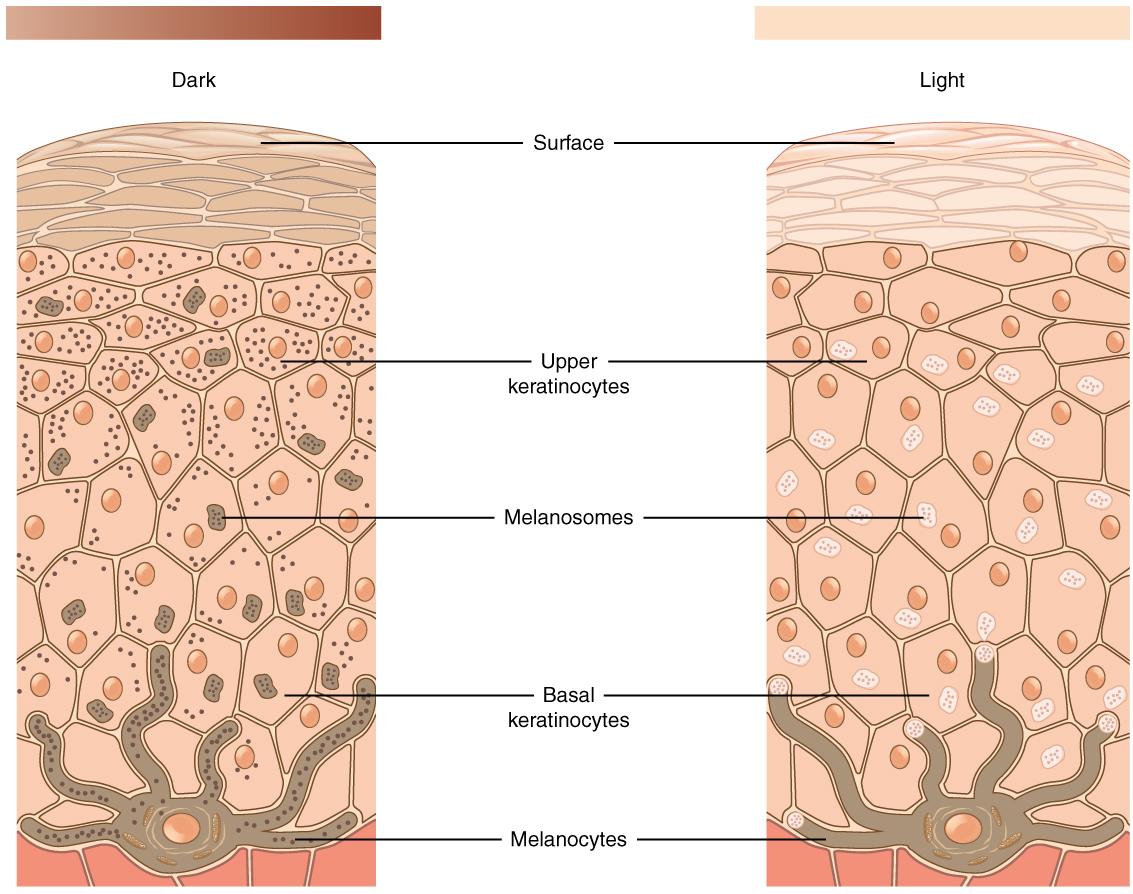 Figure \(\PageIndex{11}\): Diagram featuring the relative numbers of melanocytes and melanosomes in light and dark shades of skin tone.
Figure \(\PageIndex{11}\): Diagram featuring the relative numbers of melanocytes and melanosomes in light and dark shades of skin tone.SPECIAL TOPIC: SKIN TONE GENETIC REGULATION
The melanocortin 1 receptor (MC1R) gene acts to control which types of melanin (eumelanin or pheomelanin) are produced by melanocytes. The MC1R receptor is located on the surface of the melanocyte cells (Quillen et al. 2018). Activation of the MC1R receptors may occur through exposure to specific environmental stimuli or due to underlying genetic processes. Inactive or blocked MC1R receptors results in melanocytes producing pheomelanin. If the MC1R gene receptors are activated, then the melanocytes will produce eumelanin. Thus, individuals with activated MC1R receptors tend to have darker pigmented skin and hair than individuals with inactive or blocked receptors.
The alleles of another gene, the major facilitator, superfamily domain-containing protein 12 (MFSD12) gene, affect the expression of melanocytes in a different way than the MC1R gene. Instead of affecting the activation of melanocyte receptors, the MFSD12 alleles indirectly affect the membranes of melanocyte lysosomes (Quillen et al. 2018). The melanocyte’s lysosomes are organelles containing digestive enzymes, which ultimately correlate to varying degrees of pigmentation in humans. Variations in the membranes of the melanocyte lysosomes ultimately correlate to differing degrees of pigmentation in humans.
Ancestral MFSD12 allele variants are present in European and East Asian populations and are associated with lighter pigmentation of the skin (Crawford et al. 2017; Quillen et al. 2018). In addition, this ancestral variant is also associated with Tanzanian, San, and Ethiopian populations of Afro-Asiatic ancestry (Crawford et al. 2017; Quillen et al. 2018). In contrast, the more-derived (i.e. more recent) allele variants that are linked to darker skin tones are more commonly present in East African populations, particularly those of Nilo-Saharan descent (Crawford et al. 2017; Quillen et al. 2018). The notion that ancestral alleles of MFSD12 are associated with lighter skin pigmentation is in opposition to the commonly accepted idea that our pigmentation was likely darker throughout early human evolution (Crawford et al. 2017; Quillen et al. 2018). Due to the complexity of the human genome, MFSD12 and MC1R are but two examples of alleles affecting human skin tone. Furthermore, there is genetic evidence suggesting that certain genomic variants associated with both darker and lighter skin color have been subject to directional selection processes for as long as 600,000 years, which far exceeds the evolutionary span of Homo sapiens sapiens (Crawford et al. 2017; Quillen et al. 2018). So, evolutionary processes may lead to skin becoming more darkly pigmented as well as more lightly pigmented.
Adaptation: Melanogenesis
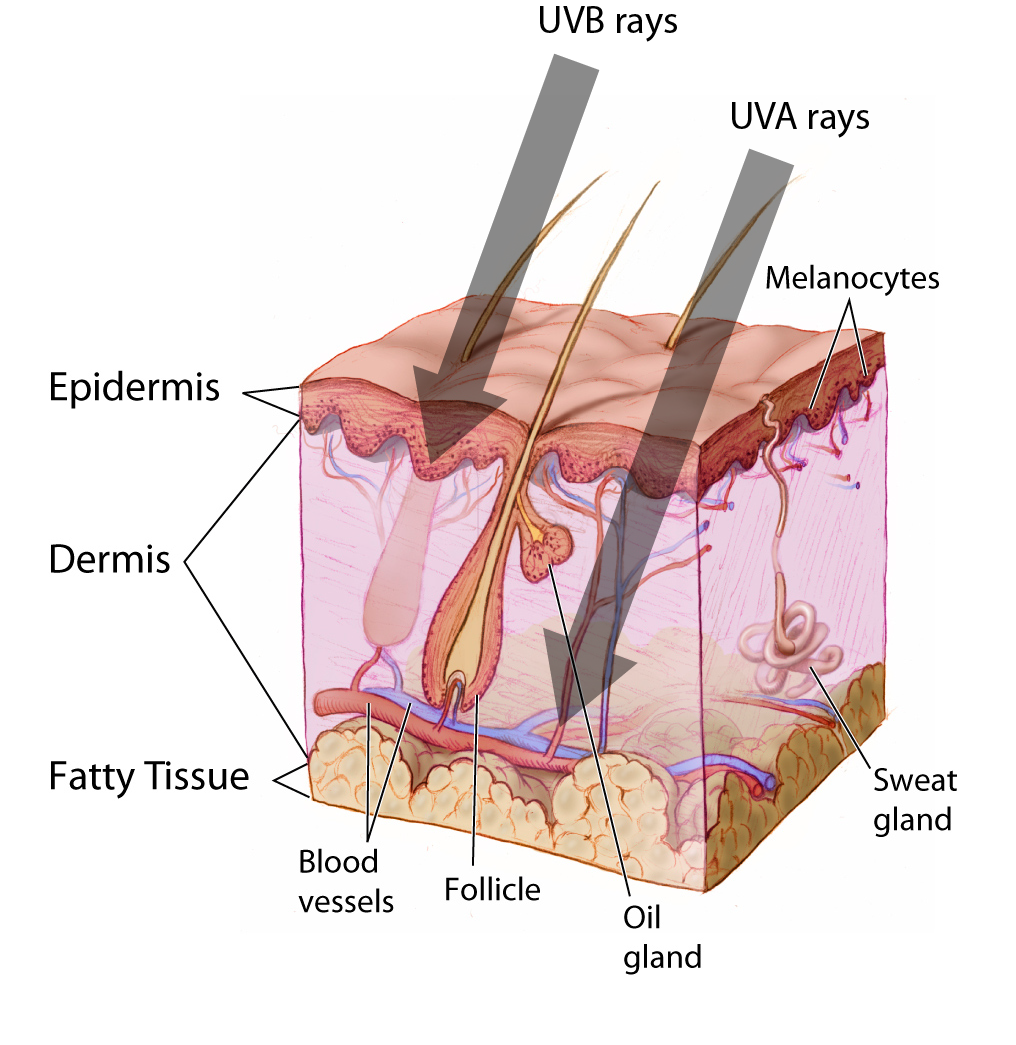 Figure \(\PageIndex{13}\): Penetration of skin layers by UVA and UVB rays.
Figure \(\PageIndex{13}\): Penetration of skin layers by UVA and UVB rays.Although all humans have approximately the same number of melanocytes within the epidermis, the production of melanin by these melanocytes varies. There are two forms of melanogenesis (the process through which melanocytes generate melanin): basal and activated. As discussed previously, the expression of eumelanin and pheomelanin by the melanocytes is genetically regulated through the expression of specific receptors (e.g., MC1R) or other melanocyte components (e.g., MFSD12). Basal melanogenesis is dependent upon an individual’s inherent genetic composition and is not influenced by external factors. Activated melanogenesis occurs in response to ultraviolet radiation (UV) exposure, specifically UV-B (short UV wave) exposure. Increased melanogenesis in response to UV-B exposure serves to provide protection to the skin’s innermost layer called the hypodermis, which lies below the epidermis and dermis (Figure 14.11). Melanin in the skin, specifically eumelanin, effectively absorbs UV-B radiation from light meaning that it will not reach the hypodermal layer. This effect is often more apparent during periods of the year when individuals tend to be outside more and the weather is warmer, which leads to those individuals donning fewer protective garments. The exposure of skin to sunlight is, of course, culturally mediated with some cultures encouraging the covering of skin at all times.
As previously noted, hemoglobin is an iron-rich protein that binds with oxygen in the bloodstream. For individuals with lighter-colored skin, blood vessels near the surface of the skin and the hemoglobin contained within those vessels is more apparent than in individuals with darker skin. The visible presence of hemoglobin coupled with the pink-to-red tone of the pheomelanin leads to lighter-skinned individuals having a pale pink skin tone. Individuals with lighter skin more readily absorb UV radiation as their basal melanin expression is directed more toward the production of pheomelanin than eumelanin. But, why are there so many variations in skin tone in humans? To answer this question, we now turn toward an exploration of an evolutionary-based adaptation of skin tone as a function of the environment.
Adaptation: Evolutionary Basis for Skin Tone Variation
 Figure \(\PageIndex{14}\): Evolutionary basis for human skin color variation.
Figure \(\PageIndex{14}\): Evolutionary basis for human skin color variation.Skin cancer is a significant concern for many individuals with light skin tone as the cumulative exposure of the epidermis and underlying skin tissues to UV radiation may lead to the development of abnormal cells within those tissues leading to malignancies. Although darker-skinned individuals are at risk for skin cancer as well, they are less likely to develop it due to increased levels of melanin, specifically eumelanin, in their skin. Even though skin cancer is a serious health concern for some individuals, most skin cancers occur in the post-reproductive years; therefore, it is improbable that evolutionary forces favoring varying melanin expression levels are related to a selective pressure to avoid such cancers. Furthermore, if avoiding skin cancer were the primary factor driving the evolution of various skin tones, then it reasons that everyone would have the most significant expression of eumelanin possible. So, why do we have different skin tones (Figure 14.12)? The term cline refers to the continuum or spectrum of gradations (i.e., levels or degrees) from one extreme to another. With respect to skin tone, the various tonal shades occur clinally
with darker skin being more prevalent near the equator and gradually decreasing in tone (i.e., decreased melanin production) in more distant latitudes. For individuals who are indigenous to equatorial regions, the increased levels of melanin within their skin provides them with a measure of protection against both sunburn and sunstroke as the melanin is more reflective of UV radiation than hemoglobin. In cases of severe sunburn, eccrine glands are affected, resulting in an individual’s ability to sweat being compromised. As sweat is the body’s most effective means of reducing its core temperature to maintain homeostasis, damage to the eccrine glands may lead to numerous physiological issues related to heat that may ultimately result in death.
Even though avoiding severe sunburn and sunstroke is of great importance to individuals within equatorial regions, this is likely not the primary factor driving the evolutionary selection of darker skin within these regions. It has been proposed that the destruction of folic acid, which is a form of B-complex vitamin, by UV radiation may have led to the selection of darker skin in equatorial regions. For pregnant women, low levels of folic acid within the body during gestation may lead to defects in the formation of the brain and spinal cord of the fetus. This condition, which is referred to as spina bifida, often significantly reduces the infant’s chances of survival without medical intervention. In men, low levels of folic acid within the body lead to an inhibition in the production of sperm. Thus, in geographic regions with high UV radiation levels (i.e., equatorial regions), there appears to be an evolutionarily driven correlation between darker skin and the maintenance of fertility.
If darker skin tone is potentially correlated to more successful reproduction, then why do lighter shades of skin exist? One hypothesis is that there is a relationship between lighter skin tone and vitamin D synthesis within the body. When skin is exposed to the UV-B radiation waves in sunlight, a series of chemical reactions occur within the epidermis leading to the production of vitamin D3. Before the body can use vitamin D3, it must travel to the liver and then to the kidneys where it is converted into different forms of bioactive molecules. Ultimately, it is converted into the bioactive molecule calcitriol (Vukić et al. 2015). Within the human body there are numerous cell types with binding receptors for calcitriol, so it is capable of adhering to the DNA of those cells (Snoddy et al. 2016). Calcitriol serves as a regulator in cellular-replication processes within the body, including those for pancreatic, breast, colon, and kidney cells (Snoddy et al. 2016). Insufficient calcitriol is associated with an increased risk of: some forms of cancer (colon, prostate, etc.), autoimmune diseases (multiple sclerosis, lupus, type I diabetes, etc.), cardiovascular diseases, and infections (e.g., tuberculosis, influenza) (Snoddy el al. 2016; Chaplin and Jablonski 2009). Deficiencies in calcitriol production and absorption within the human body may be linked to underlying genetic factors, such as a mutation in the vitamin D receptors present in some of the body’s cells (Chaplin and Jablonski 2009). Alternatively, it may be linked to inadequate exposure to the UV-B rays necessary to stimulate calcitriol production or to a nutritional deficiency in vitamin D-rich foods. Regardless of the cause of the deficiency, individuals with a calcitriol (vitamin D3) deficiency may also be at risk for the development of certain skeletal abnormalities in addition to the previously mentioned health issues.
 Figure \(\PageIndex{15}\): Children with rickets in various developmental stages.
Figure \(\PageIndex{15}\): Children with rickets in various developmental stages.Vitamin D is required for the absorption of certain nutrients, such as calcium and phosphorus, in the small intestine. These nutrients are among those that are critical for the proper growth and maintenance of bone tissue within the body. In the absence of adequate minerals, particularly calcium, bone structure and strength will be compromised leading to the development of rickets during the growth phase. Rickets is a disease affecting children during their growth phase and is characterized by inadequately calcified bones that are softer and more flexible than normal. Individuals with rickets will develop a true bowing of their femora, which may affect their mobility (Figure 14.13). In addition, deformation of pelvic bones in women may occur as a result of rickets leading to complications with reproduction. In adults, a deficiency in vitamin D will often result in osteomalacia, which is a general softening of the bones due to inadequate mineralization. This softening is the result of impaired bone metabolism that is primarily linked to insufficient levels of bioavailable vitamin D, calcium, and phosphate. In addition, it may be linked to inadequate absorption of calcium in the bloodstream. As noted, a variety of maladies may occur due to the inadequate production or absorption of vitamin D, as well as the destruction of folate within the human body; so, from an evolutionary perspective, natural selection should favor a skin tone that is best suited to a given environment.
In general, the trend related to lighter skin pigmentation further from the equator follows a principle called . This rule states that within the same species of mammals the more heavily pigmented individuals tend to originate near the equator while lighter-pigmented members of the species will be found in regions further from the equator. Gloger’s Rule applies latitudinally; however, it does not appear to hold for certain human populations near the poles. Specifically, the Inuit people (Figure 14.14), who are indigenous to regions near the North Pole and currently reside in portions of Canada, Greenland, Alaska, and Denmark. The Inuit have a darker skin tone that would not be anticipated under the provisions of Gloger’s Rule. The high reflectivity of light off of snow and ice, which is common in polar regions, necessitates the darker skin tone of these individuals to prevent folic acid degradation just as it does for individuals within equatorial regions. The consumption of vitamin D–rich foods, such as raw fish, permits the Inuit to reside at high latitudes with darker skin tone while preventing rickets.
 Figure \(\PageIndex{15}\): Inuit family, 1917.
Figure \(\PageIndex{15}\): Inuit family, 1917.Genome studies have identified a number of genes (TYR, OCA2/HERC2, TYRP1, SLC45A2, HPS6i, etc.) related to the expression of melanin and pigmentation presentation in humans. Compared to the exceptionally large number of genes within the human genome, those regulating the expression of melanin are relatively few and appear on distinct loci. The genes at these loci are generally pleiotropic in nature, so there is a relatively predictable patterning in skin, hair, and eye color combinations (Sturm and Duffy 2012). For example, some populations that are indigenous to higher latitude regions tend to have lighter skin, hair, and eye color than their counterparts from equatorial regions. Still, since the genes affecting skin, hair, and eye color are actually independent, it is possible that variations may produce many phenotypic combinations. Turning again to our example of individuals indigenous to higher latitudes, it is theoretically possible to encounter an individual with dark hair, light-toned skin, and blue eyes within this region due to the variability of phenotypic combinations.
Adaptation: Shape and Size Variations
In addition to natural selection playing a role in the determination of melanin expression related to skin tone, which is correlated to the environment, it plays a significant role in the determination of the shape and size of the human body. As previously discussed, the most significant thermodynamic mechanism of heat loss from the body is radiation. At temperatures below 20℃ (68℉), the human body loses around 65% of its heat to radiative processes; however, the efficiency of radiation as a means of heat reduction is correlated to the overall body shape and size of the individual. There is a direct correlation between the ratio of an object’s surface area to mass and the amount of heat that may be lost through radiation. For example, two metal objects of identical composition and mass are heated to the same temperature. One object is a cube and the other is a sphere. Which object will cool the fastest? Geometrically, a sphere has the smallest surface area per unit mass of any three-dimensional object, so the sphere will cool more slowly than the cube. In other words, the smaller the ratio of the surface area to mass an object has, the more it will retain heat. With respect to the cube in our example, mass increases by the cube, but surface area may increase only by the square, so size will affect the mass to surface area ratio. This, in general, holds true for humans, as well.
In regions where temperatures are consistently cold, the body shape and size of the individuals who are indigenous to the area tend to be more compact. These individuals have a relatively higher body mass to surface area (i.e., skin) than their counterparts from equatorial regions where the average temperatures are considerably warmer. Individuals from hot climates, such as the Fulani (Figure 14.15a) of West Africa, have limbs that are considerably longer than those of individuals from cold climates, such as the Inuit of Greenland (Figure 14.15b). Evolutionarily, the longer limbs of individuals from equatorial regions (e.g., the Fulani) provide a greater surface area (i.e., lower body mass to surface area ratio) for the dissipation of heat through radiative processes. In contrast, the relatively short limbs of Arctic-dwelling people, such as the Inuit, allows for the retention of heat as there is a decreased surface area through which heat may radiate away from the body.
 Figure \(\PageIndex{16}\): The Fula people of Burkina Faso (pictured here in 1974) are from a tropical environment where the rapid dispersal of heat is necessary to maintain homeostasis.
Figure \(\PageIndex{16}\): The Fula people of Burkina Faso (pictured here in 1974) are from a tropical environment where the rapid dispersal of heat is necessary to maintain homeostasis. Figure \(\PageIndex{17}\): These Inuit people from Greenland live in an arctic environment where the conservation of heat in the body’s core is of critical importance.
Figure \(\PageIndex{17}\): These Inuit people from Greenland live in an arctic environment where the conservation of heat in the body’s core is of critical importance.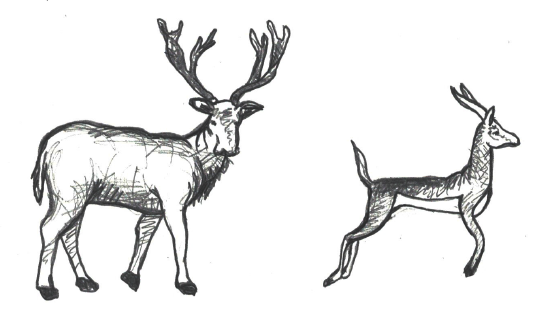 Figure \(\PageIndex{18}\): These organisms are representative of Bergmann’s rule. The animal on the left depicts an ungulate from a cooler environment with increased body weight and decreased surface area, compared to the slender ungulate on the right.
Figure \(\PageIndex{18}\): These organisms are representative of Bergmann’s rule. The animal on the left depicts an ungulate from a cooler environment with increased body weight and decreased surface area, compared to the slender ungulate on the right.As described above, there are certain trends related to the general shape and size of human bodies in relation to the thermal conditions. To better describe these trends, we turn to a couple of general principles that are applicable to a variety of species beyond humans. predicts that as average environmental temperature decreases, populations are expected to exhibit an increase in weight and a decrease in surface area (Figure 14.16a). Also, within the same species of homeothermic animals, the relative length of projecting body parts (e.g., nose, ears, and limbs) increases in relation to the average environmental temperature (Figure 14.16b). This principle, referred to as , notes that longer, thinner limbs are advantageous for the radiation of excess heat in hot environments and shorter, stockier limbs assist with the preservation of body heat in cold climates. A measure of the crural index (crural index=tibia length [divided by] femur length) of individuals from various human populations provides support for Allen’s Rule since this value is lower in individuals from colder climates than it is for those from hot climates. The crural indices for human populations varies directly with temperature, so individuals with higher crural index values are generally from regions with a warmer average environmental temperature. Conversely, the crural indices are lower for individuals from regions where there are colder average temperatures.
 Figure \(\PageIndex{19}\): These animals are representative of Allen’s rule. Note the shorter limbs and ears of the rabbit on the left that you might find in cold temperatures. Note the length of the ears on the rabbit on the right that you might find in a warm climate. Rabbits do not sweat like humans, heat is dissipated primarily through their ears.
Figure \(\PageIndex{19}\): These animals are representative of Allen’s rule. Note the shorter limbs and ears of the rabbit on the left that you might find in cold temperatures. Note the length of the ears on the rabbit on the right that you might find in a warm climate. Rabbits do not sweat like humans, heat is dissipated primarily through their ears.Nasal shape and size (Figure 14.17) is another physiological feature that is affected by way of an individual’s ancestors’ environments. The selective role of climate in determining human nasal variation is typically approached by dividing climates into four adaptive zones: hot-dry, hot-wet, cold-dry, and cold-wet (Maddux et al. 2016). One of the principal roles of the nasal cavity is to condition (i.e., warm and humidify) ambient air prior to its reaching the lungs. Given that function of the nasal cavity, it is anticipated that different nasal shapes and sizes will be related to varying environments. In cold-dry climates, an individual’s nasal cavity must provide humidification and warmth to the dry air when breathing in through the nose (Noback et al. 2011). Also, in that type of climate, the nasal cavity must conserve moisture and minimize heat loss during when the individual exhales through the nose (Noback et al. 2011). From a physiological stress perspective, this is a stressful event.
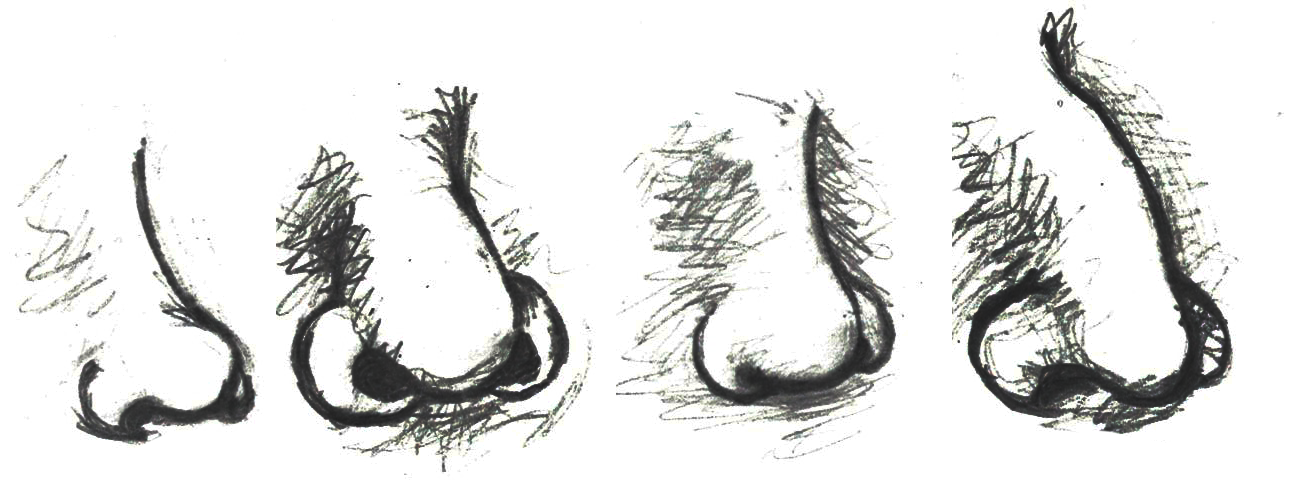 Figure \(\PageIndex{20}\): Human nasal morphological variation as influenced by four major climate-based adaptive zones: hot-dry, hot-wet, cold-dry, and cold-wet. Note that images are presented left-to-right in relation to the climate-based adaptive zones, respectively.
Figure \(\PageIndex{20}\): Human nasal morphological variation as influenced by four major climate-based adaptive zones: hot-dry, hot-wet, cold-dry, and cold-wet. Note that images are presented left-to-right in relation to the climate-based adaptive zones, respectively.Conversely, in hot-wet environments, there is no need for the nasal cavity to provide additional moisture to the inhaled air nor is there a need to warm the air or to preserve heat within the nasal cavity (Noback et al. 2011). So, in hot-wet climates, the body is under less physiological stress related to the inhalation of ambient air than in cold-dry climates. As with most human morphological elements, the shape and size of the nasal cavity occurs along a cline. Due to the environmental stressors of cold-dry environments requiring the humidification and warming of air through the nasal cavity, individuals indigenous to such environments tend to have taller (longer) noses with a reduced nasal entrance (nostril opening) size (Noback et al. 2011). This general shape is referred to as leptorrhine, and it allows for a larger surface area within the nasal cavity itself for the air to be warmed and humidified prior to entering the lungs (Maddux et al. 2016). In addition, the relatively small nasal entrance of leptorrhine noses serves as a means of conserving moisture and heat (Noback et al. 2011). Individuals indigenous to hot-wet climates tend to have platyrrhine nasal shapes, which are shorter with broader nasal entrances (Maddux et al. 2016). Since individuals in hot-wet climates do not need to humidify and warm the air entering the nose, their nasal tract is shorter and the nasal entrance wider to permit the effective cooling of the nasal cavity during respiratory processes.
Adaptation: Infectious Disease
Throughout our evolutionary journey, humans have been exposed to numerous infectious diseases. In the following section, we will explore some of the evolutionary-based adaptations that have occurred in certain populations in response to the stressors presented by select infectious diseases. One of the primary examples of natural selection processes acting on the human genome in response to the presence of an infectious disease is the case of the relationship between the sickle-cell anemia trait and malaria.
Malaria is a zoonotic disease (type of infectious disease naturally transmitted between animals and humans; covered in more detail in Chapter 16: Human Biology and Health) caused by the spread of the parasitic protozoa from the genus Plasmodium (Figure 14.18). These unicellular, eukaryotic protozoa are transmitted through the bite of a female Anopheles mosquito. During the bite process, the protozoan parasites that are present within an infected mosquito’s saliva will enter the bloodstream of the individual where they will be transported to the liver. Within the liver, the parasites multiply and will eventually be released into the bloodstream where they will infect erythrocytes. Once inside the erythrocytes, the parasites will reproduce until they exceed the cell’s storage capacity, causing it to burst and release the parasites into the bloodstream once again. This replication cycle will continue as long as there are viable erythrocytes within the host to infect.
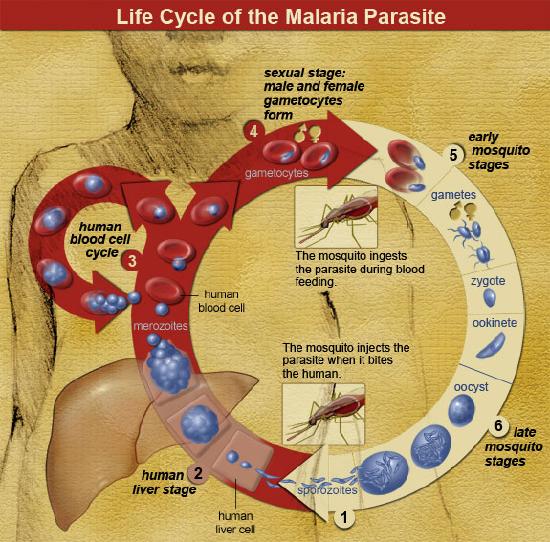 Figure \(\PageIndex{21}\): Life cycle of the malaria parasite.
Figure \(\PageIndex{21}\): Life cycle of the malaria parasite.General complications from malaria infections include: enlargement of the spleen (due to destruction of infected erythrocytes), lower number of thrombocytes (also called platelets, required for coagulation/clotting of blood), high levels of bilirubin (a byproduct of hemoglobin breakdown in the liver) in the blood, jaundice (yellowing of the skin and eyes due to increased blood bilirubin levels), fever, vomiting, retinal (eye) damage, and convulsions (seizures). According to the World Health Organization, in 2016 there were 445,000 deaths from malaria globally with the highest percentage of those deaths occurring in Africa (91%) and Southeast Asia (6%) (World Health Organization 2017). In sub-Saharan Africa, where incidents of malaria are the highest in the world, 125 million pregnancies are affected by malaria, resulting in 200,000 infant deaths (Hartman et al. 2013). Pregnant women who become infected during the gestational process are more likely to have low-birthweight infants due to prematurity or growth restriction inside the uterus (Hartman et al. 2013). After birth, infants born to malaria-infected mothers are more likely to develop infantile anemia (low red blood cell counts), a malaria infection that is not related to the maternal malarial infection, and they are more likely to die than infants born to non-malaria-infected mothers (Hartman et al. 2013).
For children and adolescents whose brains are still developing, there is a risk of cognitive (intellectual) impairment associated with some forms of malaria infections (Fernando et al. 2010). Given the relatively high rates of morbidity (disease) and mortality (number of deaths) associated with malaria, it leads to reason that this disease may have served as a selective pressure during human evolution. Support for natural selection related to malaria resistance is related to genetic mutations associated with sickle cell, thalassemia, glucose-6-phosphate dehydrogenase (G6PD) deficiency, and the absence of certain antigens (molecules capable of inducing an immune response from the host) on erythrocytes. For the purposes of this text, we will focus our discussion on the relationship between sickle cell disease and malaria.
Sickle cell disease is a group of genetically inherited blood disorders characterized by an abnormality in the shape of the hemoglobin within erythrocytes. It is important to note that there are multiple variants of hemoglobin, including, but not limited to: A, D, C, E, F, H, S, Barts, Portland, Hope, Pisa, and Hopkins. Each of these variants of hemoglobin may result in various conditions within the body; however, for the following explanation we will focus solely on variants A and S.
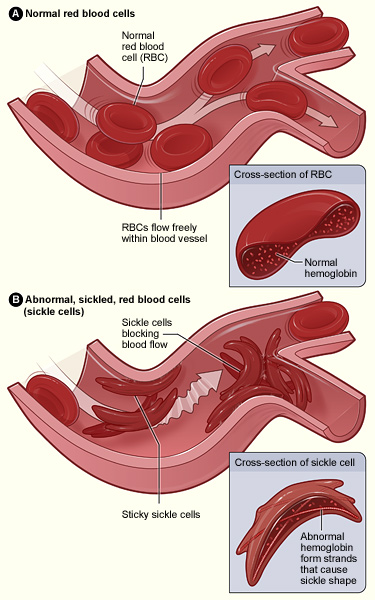 Figure \(\PageIndex{22}\): Normal and sickled erythrocytes.
Figure \(\PageIndex{22}\): Normal and sickled erythrocytes.Individuals who inherit a mutated gene (hemoglobin with a sickled erythrocyte variety, HbS) on chromosome 11 from both parents will develop sickle cell anemia, which is the most severe form of the sickle cell disease family (Figure 14.19). The genotype of an individual with sickle cell anemia is HbSS; whereas, an individual without sickle cell alleles has a genotype of HbAA representing two normal adult hemoglobin type A variants. Manifestations of sickle cell anemia (HbSS) range from mild to severe with some of the more common symptoms being: anemia, blood clots, organ failure, chest pain, fever, and low blood oxygen levels. In high-income countries with advanced medical care, the median life expectancy of an HbSS individual is around 60 years; however, in low-income countries where advanced medical care is scarce, as many as 90% of children with sickle cell disease perish before the age of five (Longo et al. 2017).
Considering that advanced medical care was not available during much of human evolutionary history, it stands to reason that the majority of individuals with the HbSS genotype died before the age of reproduction. If that is the case though, why do we still have the HbS variant present in modern populations? As covered earlier in this textbook, the genotype of an individual is composed of genes from both biological parents. In the case of an individual with an HbSS genotype, the sickle cell allele (HbS) was inherited from each of the parents. For individuals with the heterozygous genotype of HbSA, they have inherited both a sickle cell allele (HbS) and a normal hemoglobin allele (HbA). Heterozygous (HbSA) individuals who reside in regions where malaria is endemic may have a selective advantage. They will experience a sickling of some, but not all, of their erythrocytes. Unlike an individual with the HbSS genotype, someone with HbSA may experience some of the symptoms listed above; however, they are generally less severe.
As noted earlier, the mechanism through which Plasmodium protozoan parasites replicate involves human erythrocyte cells. However, due to their sickled shape, as well as the presence of an abnormally shaped protein within the cell, the parasites are unable to replicate effectively in the erythrocyte cells coded for by the HbS allele (Cyrklaff et al. 2011). An individual who has an HbSA genotype and an active malaria infection will become ill with the disease to a lesser extent than someone with an HbAA genotype. Although normal erythrocytes (regulated by the HbA allele) allow for the replication of the parasite, the parasites will not be able to replicate in HbS erythrocytes of the heterozygote. So, individuals with the HbSA genotype are more likely to survive a malaria infection than an individual who is HbAA. Although individuals with the HbSA genotype may endure some physiological complications related to the sickling of some of their erythrocytes, their morbidity and mortality rates are lower than they are for HbSS members of the population. The majority of individuals who are heterozygous or homozygous for the HbS trait have ancestors who originated in sub-Saharan Africa, India, Saudi Arabia, regions in South and Central America, the Caribbean, and the Mediterranean (Turkey, Greece, and Italy) (Centers for Disease Control and Prevention 2017) (Figure 14.20).
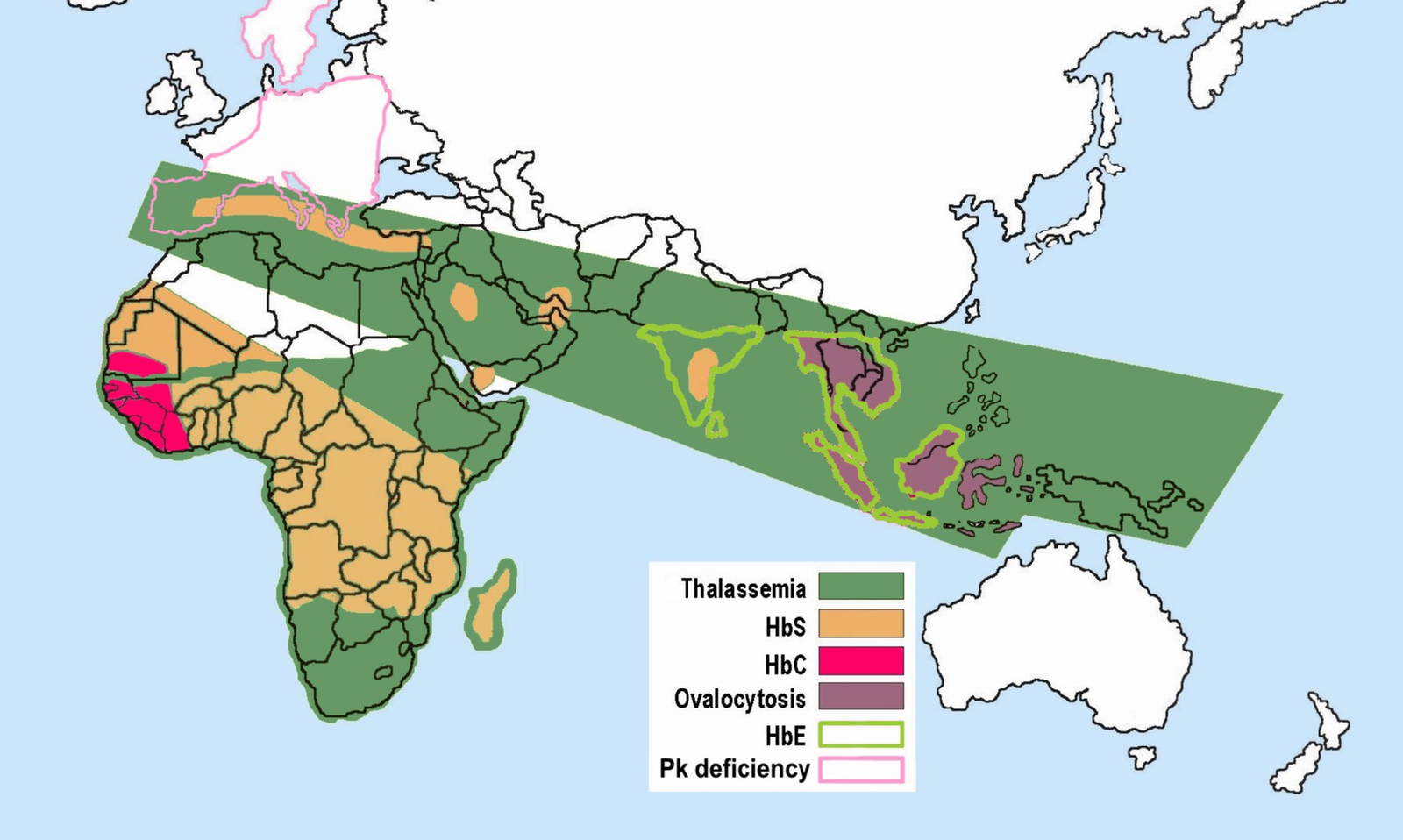 Figure \(\PageIndex{23}\): Distribution of sickle cell and associated erythrocytic abnormalities for Africa and Asia.
Figure \(\PageIndex{23}\): Distribution of sickle cell and associated erythrocytic abnormalities for Africa and Asia.With respect to the history of these regions, during the early phases of settlement horticulture was the primary method of crop cultivation. Typically performed on a small scale, horticulture is based on manual labor and relatively simple hand tools rather than the use of draft animals or irrigation technologies. Common in horticulture is swidden, or the cutting and burning of plants in woodland and grassland regions. The swidden is the prepared field that results following a slash-and-burn episode. This practice fundamentally alters the soil chemistry, removes plants that provide shade, and increases the areas where water may pool. This anthropogenically altered landscape provides the perfect breeding ground for the Anopheles mosquito, as it prefers warm, stagnant pools of water (Figure 14.21).
 Figure \(\PageIndex{24}\): The effects of human horticultural activities on the balancing selection of populations in relation to sickle cell disease genotype variants.
Figure \(\PageIndex{24}\): The effects of human horticultural activities on the balancing selection of populations in relation to sickle cell disease genotype variants.Although swidden agriculture was historically practiced across the globe, it became most problematic in the regions where the Anopheles mosquito is endemic. These areas have the highest incidence rates of malaria infection. Over time, the presence of the Anopheles mosquito and the Plasmodium parasite that it transmitted acted as a selective pressure, particularly in regions where swidden agricultural practices were common, toward the selection of individuals with some modicum of resistance against the infection. In these regions, HbSS and HbSA individuals would have been more likely to survive and reproduce successfully. Although individuals and populations are far more mobile now than they have been throughout much of history, there are still regions where we can see higher rates of malaria infection as well as greater numbers of individuals with the HbS erythrocyte variant. The relationship between malaria and the selective pressure for the HbS variant is one of the most prominent examples of natural selection in the human species within recent evolutionary history.
Adaptation: Lactase Persistence
With the case of sickled erythrocytes and their resistance to infection by malaria parasites, there is strong support for a cause-and-effect-style relationship linked to natural selection. Although somewhat less apparent, there is a correlation between lactase persistence and environmental challenges. Lactase-phlorizin hydrolase (LPH) is an enzyme that is primarily produced in the small intestine and permits the proper digestion of lactose, a disaccharide (composed of two simple sugars: glucose and galactose) found in the milk of mammals. Most humans will experience a decrease in the expression of LPH following weaning, leading to an inability to properly digest lactose. Generally, LPH production decreases between the ages of two and five and is completely absent by the age of nine (Dzialanski et al. 2016). For these individuals, the ingestion of lactose may lead to a wide variety of gastrointestinal ailments including abdominal bloating, increased gas, and diarrhea. Although the bloating and gas are unpleasant, the diarrhea caused by a failure to properly digest lactose can be life-threatening if severe enough due to the dehydration it can cause. Some humans, however, are able to produce LPH far beyond the weaning period.
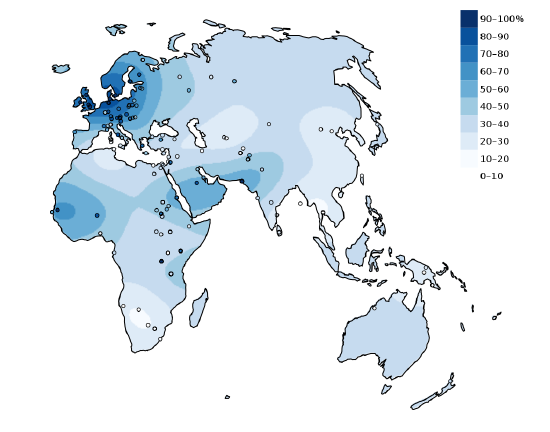 Figure \(\PageIndex{25}\): Interpolated map depicting the percentage of adults with the lactase persistence genotype in indigenous populations of the Old World. Circles denote sample locations.
Figure \(\PageIndex{25}\): Interpolated map depicting the percentage of adults with the lactase persistence genotype in indigenous populations of the Old World. Circles denote sample locations.Individuals who continue to produce LPH have what is referred to as the lactase persistence trait. The lactase persistence trait is encoded for a gene called LCT, which is located on human chromosome 2 (Ranciaro et al. 2014; see also Chapter 3). From an evolutionary and historical perspective, this trait is most commonly linked to cultures that have practiced cattle domestication (Figure 14.22). For individuals in those cultures, the continued expression of LPH may have provided a selective advantage. During periods of environmental stress, such as a drought, if an individual is capable of successfully digesting cow’s milk, they have a higher chance of survival than someone who suffers from diarrhea-linked dehydration due to a lack of LPH. Per Tishkoff et al. , the “frequency of lactase persistence is high in northern European populations (more than 90% in Swedes and Danes), decreases in frequency across southern Europe and the Middle East (less than 50% in Spanish, French, and pastoralist Arab populations), and is low in non-pastoralist Asian and African populations (less than 1% in Chinese, less than 5% to 20% in West African agriculturalists)” (2007: 248). Although the frequency of the lactase persistence trait is relatively low among African agriculturalists, it is high among pastoralist populations that are traditionally associated with cattle domestication, such as the Tutsi and Fulani, who have frequencies of 90% and 50%, respectively (Tishkoff et al. 2007).
Cattle domestication began around 11,000 years ago in Europe (Beja-Pereira et al. 2006) and 7,500 to 9,000 years ago in the Middle East and North Africa (Tishkoff et al. 2007). Based on human genomic studies, it is estimated that the mutation for the lactase persistence trait occurred around 2,000 to 20,000 years ago for European populations (Tishkoff et al. 2007). For African populations, the lactase persistence trait emerged approximately 1,200 to 23,000 years ago (Gerbault et al. 2011). This begs the question: Is this mutation the same for both populations? It appears that the emergence of the lactase persistence mutation in non-European populations, specifically those in East Africa (e.g., Tutsi and Fulani), is a case of convergent evolution. With convergent evolution events, a similar mutation may occur in species of different lineages through independent evolutionary processes. Based on our current understanding of the genetic mutation pathways for the lactase persistence trait in European and African populations, these mutations are not representative of a shared lineage. In other words, just because a person of European origin and a person of African origin can each digest milk due to the presence of the lactase-persistence trait in their genotypes, it does not mean that these two individuals inherited it due to shared common ancestry.
Is it possible that the convergent evolution of similar lactase-persistence traits in disparate populations is merely a product of genetic drift? Or is there evidence for natural selection? Even though 23,000 years may seem like a long time, it is but a blink of the proverbial evolutionary eye. From the perspective of human evolutionary pathways, mutations related to the LCT gene have occurred relatively recently. Similar genetic changes in multiple populations through genetic drift processes, which are relatively slow and directionless, fail to accumulate as rapidly as have lactase-persistence traits (Gerbault et al. 2011). The widespread accumulation of these traits in a relatively short period of time supports the notion that an underlying selective pressure must be driving this form of human evolution. Although to date no definitive factors have been firmly identified, it is thought that environmental pressures are likely to credit for the rapid accumulation of the lactase-persistence trait in multiple human populations through convergent evolutionary pathways.
Human Variation: Our Story Continues
From the time that the first of our species left Africa, we have had to adjust and adapt to numerous environmental challenges. The remarkable ability of human beings to maintain homeostasis through a combination of both nongenetic (adjustments) and genetic (adaptations) means has allowed us to occupy a remarkable variety of environments from high-altitude mountainous regions to the tropics near the equator. From adding piquant, pungent spices to our foods as a means of inhibiting food-borne illnesses due to bacterial growth to donning garments specially suited to local climates, behavioral adjustments have provided us with a nongenetic means of coping with obstacles to our health and well-being. Acclimatory adjustments, such as sweating when we are warm in an attempt to regulate our body temperature or experiencing increased breathing rates as a means of increasing blood oxygen levels in regions where the partial pressure of oxygen is low, have been instrumental in our survival with respect to thermal and altitudinal environmental challenges. For some individuals, developmental adjustments that were acquired during their development and growth phases (e.g., increased heart and lung capacities for individuals from high-altitude regions) provide them with a form of physiological advantage not possible for someone who ventures to such an environmentally challenging region as an adult. Genetically-mediated adaptations, such as variations in the pigmentation of our skin, have ensured our evolutionary fitness across all latitudes.
Will the human species continue to adjust and adapt to new environmental challenges in the future? If past performance is any measure of future expectations, then the human story will continue as long as we do not alter our environment to the point that the plasticity of our behavior, physiological, and morphological boundaries is exceeded. In the following chapters, you will explore additional information about our saga as a species. From the concept of race as a sociocultural construct to our epidemiological history, the nuances of evolutionary-based human variation are always present and provide the basis for understanding our history and our future as a species.