
2.2: Darwinian Theory
- Page ID
- 158719
THE TRANSMUTATION HYPOTHESIS
According to the Bible (Genesis 11), all people once spoke a single language and decided to cooperate to build a giant tower that would stretch into the heavens. For this arrogance, they are made to speak different languages and must give up building the tower. The story’s setting is generally thought to refer to the ancient ziggurats of Babylonia.
The idea that species had their beginnings in other, older, similar species.
Darwin’s term for what we now call “evolution,” in which animals and plants look different from their ancestors.
 Figure \(\PageIndex{1}\): A Beagle
Figure \(\PageIndex{1}\): A Beagle Figure \(\PageIndex{2}\): Bulldog
Figure \(\PageIndex{2}\): Bulldog Figure \(\PageIndex{3}\): Greyhound
Figure \(\PageIndex{3}\): GreyhoundThe process whereby organisms that are better adapted to their environment tend to survive longer and transmit more of their genetic characteristics to succeeding generations than do those that are less well adapted.
As defined by Edward Tylor: That complex whole which includes knowledge, belief, art, morals, law, custom, and any other capabilities and habits acquired by man as a member of society (Tylor 1871, 1).
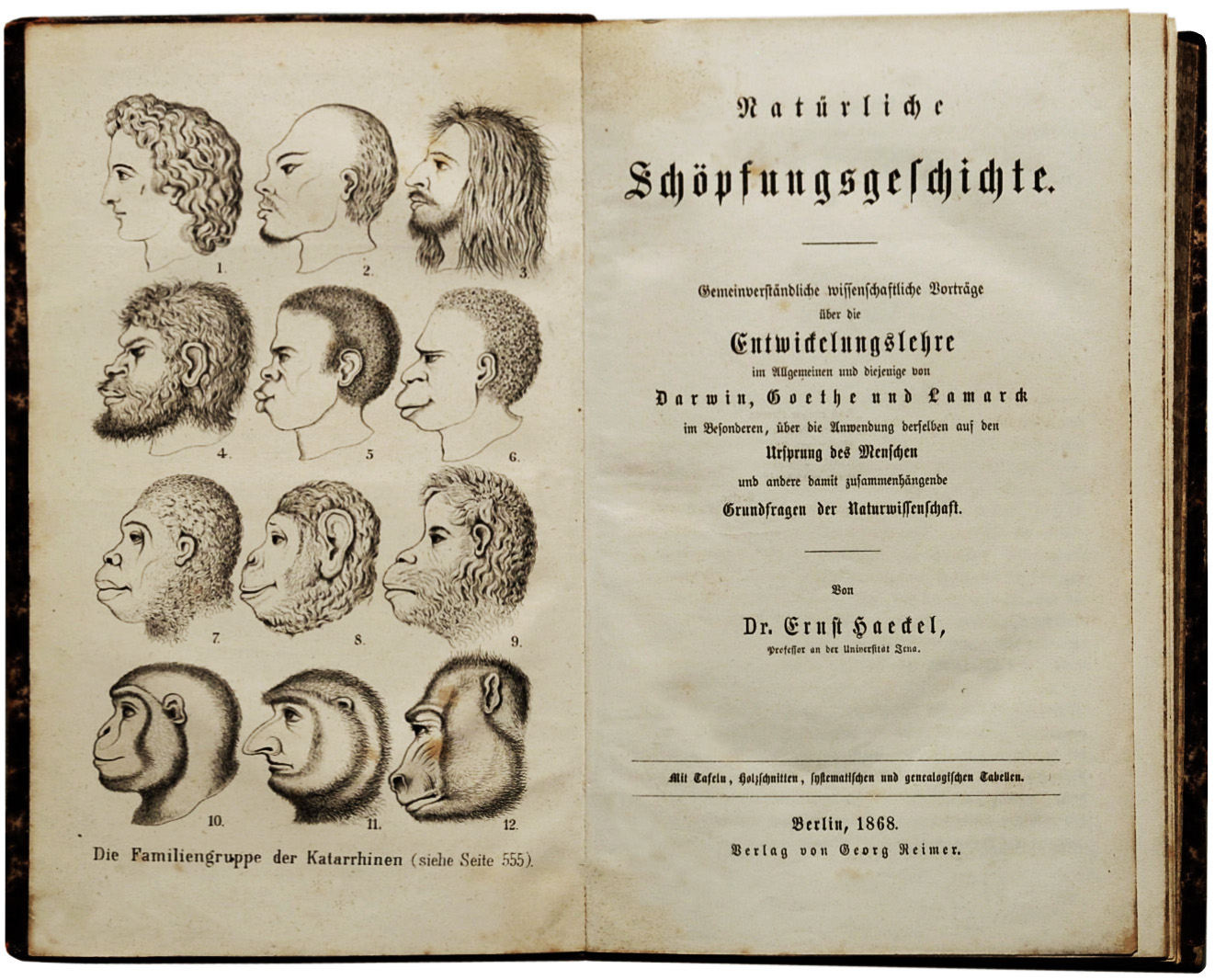 Figure \(\PageIndex{4}\): The frontispiece to Ernst Haeckel’s (1868) popular German book on Darwinism. The English translation lacked this illustration.
Figure \(\PageIndex{4}\): The frontispiece to Ernst Haeckel’s (1868) popular German book on Darwinism. The English translation lacked this illustration.POST-DARWINIAN THEORIES AND DISPUTES
The immediate theoretical weakness of Darwinism lay in its reliance upon a pool of undirected variation for nature to select from. The dominant theory of heredity at the time was known as blending inheritance, in which a child is a blend of the parents—like paint, if mom is red and dad is blue, then the child is purple (see Figure 2.2.5). The problem is that any descendants of purple child will never be as different as blue mom and red dad. You can’t recover the original blue and red from purple paint—which simply means that for people, variation is lost every generation. How can natural selection work if you lose variation every generation?
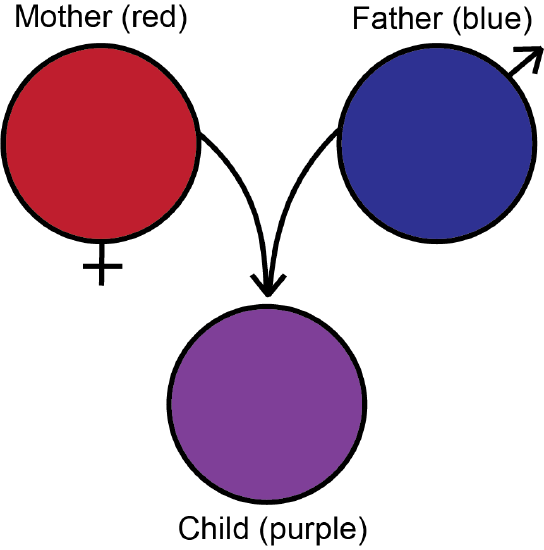
An obsolete theory where the offspring of two parents will have characteristics that are intermediate between the two parents.
Darwin fell back on a principle developed by Lamarck known as the inheritance of acquired characteristics or “use and disuse of organs.” Here, whatever attributes you develop over the course of your life—muscles, a tan, compassion, bad breath—can be stably passed on to your children, somehow. That way, variation can be reintroduced every generation, by virtue of this new pool of acquired characters. Unfortunately, an influential school of German biologists in the 1880s, led by August Weismann, had identified just two types of cells in bodies: reproductive or germ cells, and somatic or body cells. It was the germ cells that formed the next generation; the somatic cells, which form the body, comprise merely an evolutionary dead-end to aid in the transmission of the germ-line. Life could thus be seen as a continuous series of germ-cells, with adult bodies as transient receptacles grown up around them every generation. On this basis, the English writer Samuel Butler quipped that a hen is just an egg’s way of making another egg. But how, then, could information about your elbow or your cerebral cortex during the course of your life get into your germ cells? There didn’t seem to be a way, so that generation called themselves “neoDarwinians” to express their belief in natural selection minus the inheritance of acquired characteristics.
The idea that you pass on the traits that you have acquired during your lifetime, not just your genes; also known as Lamarckian inheritance.
The entire problem was rendered moot with the discovery in 1900 of Gregor Mendel’s work on heredity in peas from 35 years earlier. Mendel showed that heredity didn’t actually work like the blending of paints at all. When you isolated particular traits, you saw that offspring were not midway between their parents; rather, they were like one or the other parent. The offspring of a plant with green peas and one with yellow peas was green, not chartreuse. The offspring of a plant with wrinkled peas and one with round peas was round, not wrinkly-round. This suggested, rather, that heredity worked like interacting particles that came into new combinations but fundamentally retained their structural integrity every generation. Unlike paints, you could indeed recover the original variants under this model; variation wasn’t lost every generation.
Mendelian genetics soon created new problems for Darwinism, however. The new geneticists were focused on discrete binary states of existence, like Mendel’s peas: green/yellow, wrinkled/round, tall/short, in experimental populations. But the old Darwinian naturalists were working with quantitative variations in real populations—many of them intermediate, not extreme, in form. So, the Mendelians had a robust theory of heredity that had difficulty explaining natural patterns of variation, and the Darwinians had a robust theory of biological change that had difficulty accommodating discontinuous variation. One solution might be to reconceptualize all variation as fundamentally binary; the American geneticist Charles Davenport, for example, argued with considerable success that there were two kinds of people—smart and stupid—and that the stupid people simply had the allele for “feeblemindedness.” This actually had a major and regrettable impact on American science and social policy in the 1920s.
A genetic variant. A non-identical DNA sequence found in the same gene location on a homologous chromosome, or gene copy, that codes for the same trait but produces a different phenotype.
A better solution came with the invention of population genetics, in works published around 1930 by the British geneticists Ronald Fisher and J. B. S. Haldane and the American geneticist Sewall Wright. In this model, a gene has small but cumulative effects. If we reduce a body to its genetic composition or genotype, and we reduce a species to its cumulative genetic composition, or gene pool, we can mathematically model the ways in which the gene pool can be transformed. There are rather few ways to accomplish it, and each has characteristic and predictable effects.
Segment of DNA that contains protein-coding information and various regulatory (e.g., promoter) and noncoding (e.g., introns) regions.
The entire set of genes of an individual organism; the combination of two alleles that code for or are associated with the same gene.
The entire collection of genetic material in a breeding community that can be passed on from one generation to the next.
 Figure \(\PageIndex{6}\): George Gaylord Simpson (1983). Photo courtesy of Jonathan Marks.
Figure \(\PageIndex{6}\): George Gaylord Simpson (1983). Photo courtesy of Jonathan Marks.This became the first part of the Synthetic Theory of Evolution, the extension of Mendelian genetics to population genetics and the formal mathematical study of how gene pools may be transformed through time. The second part involved the study of how species diversify in addition to simply changing, and it entailed integrating speciation and geography in the story of how animal species have come to be. The primary scholars involved were the Russian-American fruit fly geneticist Theodosius Dobzhansky, the German-American ornithologist Ernst Mayr, and the American paleontologist George Gaylord Simpson (see Figure 2.2.6).
Explanation of the evolution of life in terms of genetic changes occurring in the population that leads to the formation of new species.
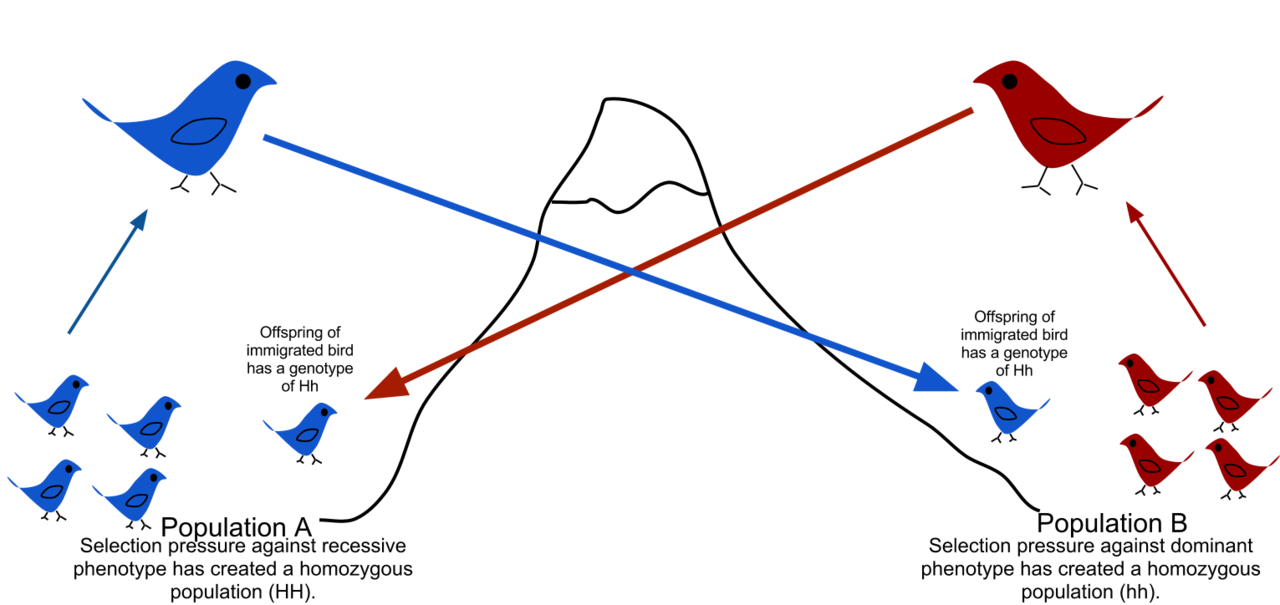
By the 1960s, biologists had a robust theory to explain the history of life. Genetic or genotypic changes (known to be encoded in molecules of DNA) cause changes in the physical appearance or phenotype. The environment sorts out these changes, and their proportion within a species rises or falls with the nature and stringency of the environment. Selection could now be reduced to the favoring of certain genotypes over alternatives, which can make populations genetically and adaptively different from one another. Genetic drift, or stochastic (random) changes to the gene pool, makes populations genetically different from one another nonadaptively—that is to say, in ways that don’t track the environment. The genetic contact of populations, or gene flow, makes populations more similar to one another. Disrupting gene flow acts to divide gene pools, which is in turn stabilized by the development of reproductive barriers between the populations. These processes can be directly studied within living species and can be extrapolated and can adequately explain the differences we find among species.
Observable physical or biochemical traits of an organism that are produced through the interaction of genotype and environment.
Random changes in allele frequencies within a population from one generation to the next with nonadaptive effects.
Geographical movement of genes due to the transfer of alleles from one population to another through the contact of different populations.
REFERENCES
Darwin, Charles. 1859. On the origin of species by means of natural selection, or, the preservation of favoured races in the struggle for life. London: J. Murray.
Darwin, Charles. 1869. Correspondence Project. “Letter no. 6684,” accessed on 11 July 2019, http://www.darwinproject.ac.uk/DCP-LETT-6684.
Tylor, Edward B. 1871. Primitive Culture. London: John Murray.
FIGURE ATTRIBUTIONS
Figure 2.2.1 Pet (pet-dog-animals-cute-animal-dogs-3635986/) by Somo_Photography has been designated to the public domain (CC0).
Figure 2.2.2 English bulldog (2705136) by mk817 has been designated to the public domain (CC0) .
Figure 2.2.3 ItalianGreyhound by Just chaos is used under a CC BY 2.0 license.
Figure 2.2.4 Ernst Haeckel – Natürliche Schöpfungsgeschichte, 1868 by Foto H.-P.Haack Das Foto darf gebührenfrei verwendet werden, sofern der Urheber mit “Foto H.-P.Haack” vermerkt wird. is used under a CC BY-SA 3.0 DE license.
Figure 2.2.5 Blending inheritance in color original to Explorations: An Open Invitation to Biological Anthropology by Mary Nelson is under a CC BY-NC 4.0 License.
Figure 2.2.6 George Gaylord Simpson (1983) photo courtesy of Jonathan Mark is under a CC BY-NC 4.0 License.
Figure 2.2.7 Gene Flow (2012) by Jessica Krueger, CC BY-SA 3.0, via Wikimedia Commons.