
2.3: Molecular Revolution
- Page ID
- 158721
GENETICS AND ANATOMY
 Figure \(\PageIndex{1a}\): Horse.
Figure \(\PageIndex{1a}\): Horse. Figure \(\PageIndex{1b}\): Donkey.
Figure \(\PageIndex{1b}\): Donkey. Figure \(\PageIndex{2}\): Dentition of Ramapithecus.
Figure \(\PageIndex{2}\): Dentition of Ramapithecus.An alteration to the base sequence in the DNA of an organism.
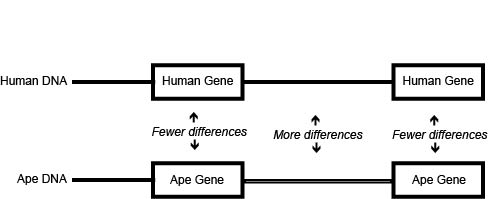 Figure \(\PageIndex{3}\): DNA comparisons yield more difference between than within genes.
Figure \(\PageIndex{3}\): DNA comparisons yield more difference between than within genes.A change in the DNA sequence that codes for amino acids in a protein sequence, but does not change the encoded amino acid.
ORGANISMAL AND MULTILEVEL EVOLUTION
By the 1980s, the acknowledgment that even though genes cause bodies, genes and bodies evolve with different rates and patterns, led to a renewed focus on how bodies change. The Evolutionary Synthesis of the 1930s–1970s had reduced organisms to their genotypes and species to their gene pools, which provided valuable insights about the processes of biological change, but it was only a first approximation. Animals are in fact reactive and adaptable beings, not passive and inert genotypes. Nor are species simply gene pools; rather, they are clusters of socially interacting and reproductively compatible organisms.
So, accepting that evolutionary change is fundamentally genetic change, how do bodies nevertheless function and evolve? And accepting that speciation is ultimately a division of the gene pool, how do groups of animals nevertheless come to see one another as potential mates or competitors for mates, as opposed to just other creatures in the environment? Are there evolutionary processes that are not explicable by population genetics? These questions were raised in the 1980s by paleontologist Stephen Jay Gould, the leading evolutionary biologist of the late 20th century, to progress beyond the reductive assumptions that had guided the earlier generation.
Gould spearheaded a movement to identify and examine higher-order processes and features of evolution that were not adequately explained by population genetics. For example, extinction, which was such a problem for biologists of the 1600s, could now be seen as playing a more complex role in the history of life than population genetics had been able to model. The crucial recognition was that there are two kinds of extinctions, each with different consequences: background extinctions and mass extinctions. Background extinctions are those that reflect the balance of nature, because in a competitive Darwinian world, some things go extinct and other things take their place. Ecologically, your species may be adapted to its niche, but if another species comes along that’s better adapted to the same niche, eventually your species will go extinct. It sucks, but it is the way of all life: you come into existence, you endure, and you pass out of existence. But mass extinctions are quite different. They reflect the wholesale disruption of nature rather than the balance of nature, such as many species from many different lineages dying off at roughly the same time—presumably as the result of some kind of rare ecological disaster. The situation may not be survival of the fittest as much as survival of the luckiest. The result, then, would be an ecological scramble among the survivors. Having made it through the worst, the survivors could now simply divide up the new ecosystem amongst themselves, since their competitors were gone. Something like this may well have happened about 65 million years ago, with mammals surviving and dinosaurs not. Something like this may be happening now, due to human expansion and environmental degradation. Note, though, that there is only a limited descriptive role here for population genetics: the phenomena we are describing are about organisms and species in ecosystems.
Another question involved the properties of species that might not be reducible to the properties of their gene pools. For example, there are upwards of 15 species of gibbons but only two of chimpanzees. Why? There are upwards of 20 species of guenons but fewer than ten of baboons. Why? Are there genes for that? It seems unlikely. Gould suggested that species, as analytic units of nature, might have properties that are not reducible to the genes in their cells. For example, characteristic rates of speciation and extinction might be emergent properties of their ecologies and histories, and not properties of the genes. Consistent biases of speciation rates might well produce patterns of macroevolutionary diversity that are difficult to explain genetically and that need to be understood ecologically. Gould called such biases in speciation rates species selection—a higher-order process that invokes competition between species, in addition to the classic Darwinian competition between individuals.
A postulated evolutionary process which suggests that selection acts on an entire species population, rather than individuals.
One of Gould’s most important studies involved the very nature of species. In the classical view, a species is continually adapting to its environment until it changes so much that it is a different species (phyletic gradualism) than it was at the beginning of this sentence (Eldredge and Gould 1972). That implies that the species is a fundamentally unstable entity through time, continuously changing to fit in. But suppose, argued Gould along with paleontologist Niles Eldredge, a species is more fundamentally stable through time and only really adapts as it is being founded? Then we might expect to find in the fossil record long equilibrium periods—a few million years or so—in which species don’t seem to change much, punctuated by relatively brief periods in which they change a bit and then stabilize again as new species. They called this idea punctuated equilibria, and it helps to explain certain features of the fossil record, notably the existence of small anatomical “gaps” between closely related fossil forms (see Figure 2.4.4). Its significance, once again, lies in the fact that although it incorporates genetics, it is not really a theory of genetics but a theory of groups of bodies in deep time.
The theory that evolution occurs slowly and uniformly over long periods of time; speciation is a gradual process.
The theory that evolution occurs rapidly over brief periods of change and speciation, followed by long periods of status or equilibrium.
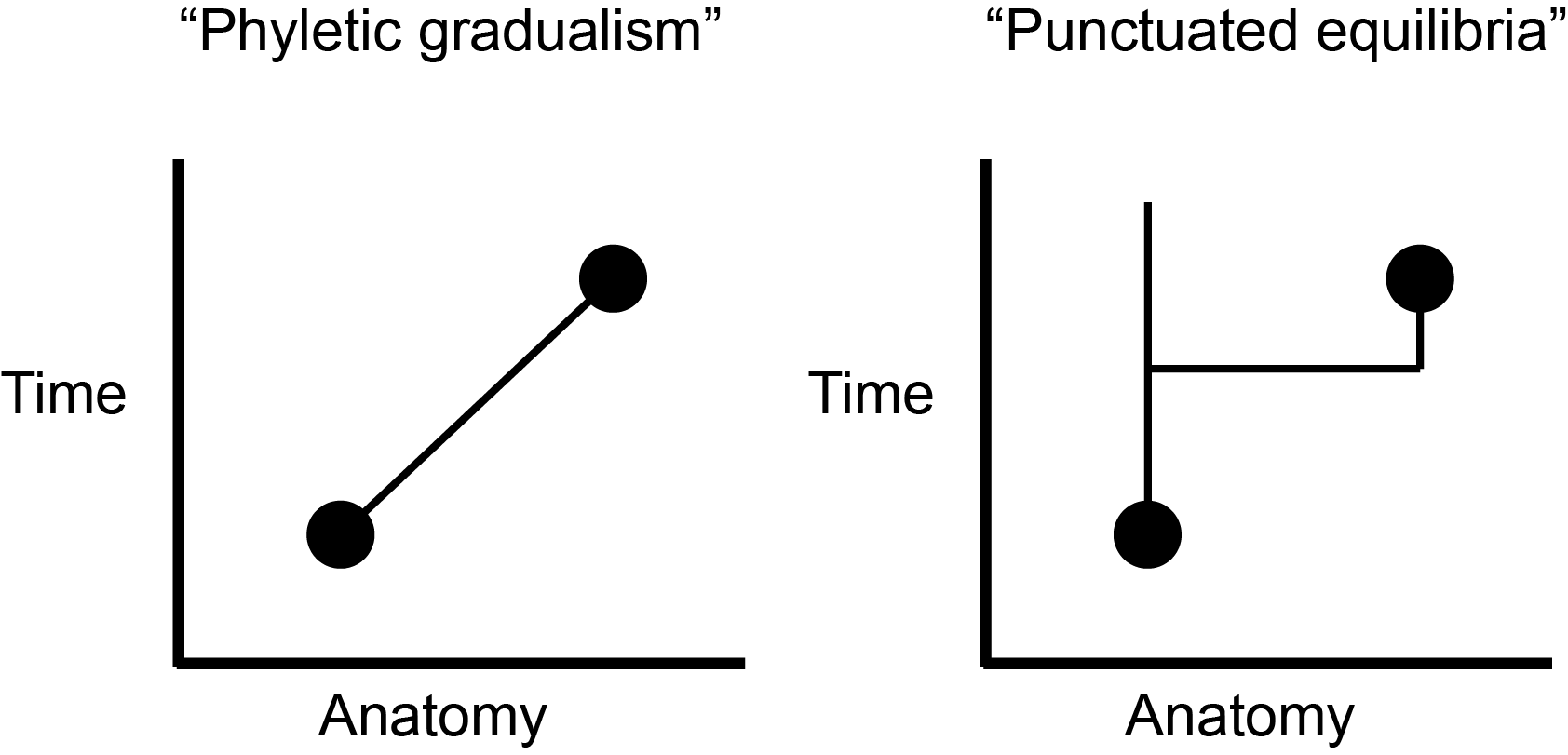 Figure \(\PageIndex{4}\): Different ways of conceptualizing the evolutionary relationship between an earlier and a later species.
Figure \(\PageIndex{4}\): Different ways of conceptualizing the evolutionary relationship between an earlier and a later species.In response to the call for a theory of the evolution of form, the field of evo-devo—the intersection of evolutionary and developmental biology—arose. The central focus here is on how changes in form and shape arise. An embryo matures by the stimulation of certain cells to divide, forming growth fields. The interactions and relationships among these growth fields generate the structures of the body. The genes that regulate these growth fields turn out to be very highly conserved across the animal kingdom. This is because they repeatedly turn on and off the most basic genes guiding the animal’s development, and thus any changes to them would be catastrophic. Indeed, these genes were first identified by producing a bizarre mutant fruit fly that grew a pair of legs where its antennae were supposed to be.
The study of the origin of form; a contraction of “evolutionary developmental biology.”
Certain genetic changes can alter the fates of cells and the body parts that they build; meanwhile, other genetic changes can simply affect the rates at which neighboring groups of cells grow and divide, thus producing physical bumps or dents in the developing body. The result of altering the relationships among these fields of cellular proliferation in the growing embryo is allometry, or the differential growth of body parts. As an animal gets larger—either over the course of its life or over the course of macroevolution—it often has to change shape in order to live at a different size. Many important physiological functions depend on properties of geometric area: the strength of a bone, for example, is proportional to its cross-sectional area. But area is a two-dimensional quality, while growing takes place in three dimensions—as an increase in mass or volume. As an animal expands, its bones necessarily weaken, because volume expands faster than area does. Consequently, a bigger animal has more stress on its bones than a smaller animal does and must evolve bones even thicker than they would be by simply scaling the animal up proportionally. In other words, if you expand a mouse to the size of an elephant, it will nevertheless still have much thinner bones than the elephant does. But those giant mouse bones will unfortunately not be adequate to the task. Thus, a giant mouse would have to change aspects of its form to maintain function at a larger size (see Figure 2.4.5).
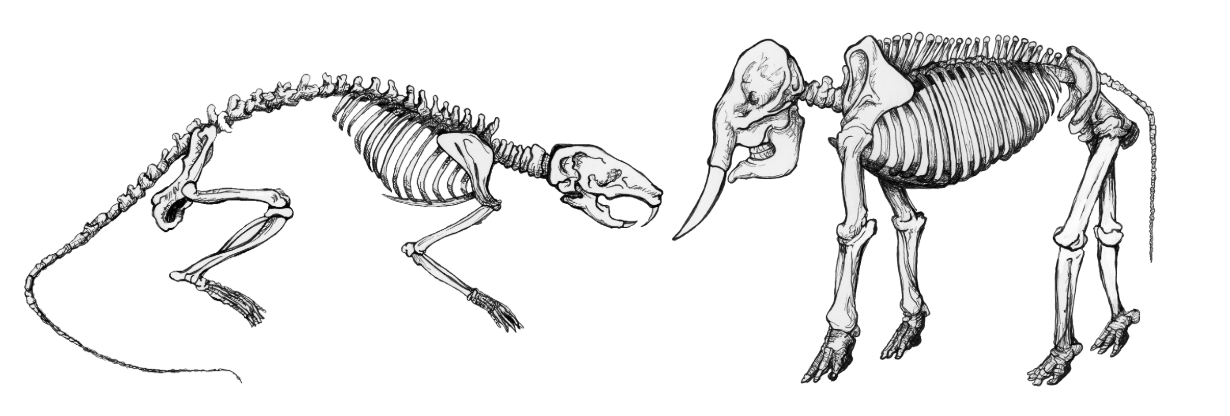 Figure \(\PageIndex{5}\): Mouse (left) and elephant skeletons (right). Notice the elephant’s bones are more robust when the two animals are the same size.
Figure \(\PageIndex{5}\): Mouse (left) and elephant skeletons (right). Notice the elephant’s bones are more robust when the two animals are the same size.Physiologically, we would like to know how the body “knows” when to turn on and off the genes that regulate growth to produce a normal animal. Evolutionarily, we would like to know how the body “learns” to alter the genetic on/off switch (or the genetic “slow down/speed up” switch) to produce an animal that looks different. Moreover, since organisms differ from one another, we would like to know how the developing body distinguishes a range of normal variation from abnormal, pathological variation. And finally, how does abnormal variation eventually become normal in a descendant species?
Gould here invoked the work of a British geneticist named Conrad H. Waddington, who thought about genetics less reductively than his colleagues. Without isolating specific DNA sites and analyzing their function, Waddington instead studied the inheritance of an organism’s reactivity—its ability to adapt to the circumstances of its life. In a famous experiment, he grew fruit fly eggs in an atmosphere containing ether. Most died, but a few survived somehow by developing a weird physical feature: a second thorax, with a second pair of wings. Waddington bred these flies and soon developed a stable line of flies who would reliably develop a second thorax when grown in ether. Then he began to lower the concentration of ether, while continuing to selectively breed the flies that developed the strange appearance. Eventually, he had a line of flies that would stably develop the “bithorax” phenotype even when there was no ether; it had become the “new normal.” The flies had genetically assimilated the bithorax condition.
Waddington was thus able to mimic the inheritance of acquired characteristics: what had been a trait stimulated by ether a few generations ago was now a normal part of the development of the descendants. Waddington recognized that he had performed a selection experiment on genetic variants, yet he had not selected for particular traits but, rather, for the physiological tendency to develop particular traits when appropriately stimulated. He called that tendency plasticity and its converse, the tendency to stay the same even under weird environmental circumstances, canalization. Waddington had initially selected for plasticity, the tendency to develop the bithorax phenotype under weird conditions, and then, later, for canalization, the developmental normalization of that weird physical trait. Although Waddington had high stature in the community of geneticists, evolutionary biologists of the 1950s and 1960s regarded him with suspicion because he was not working within the standard mindset of reductionism, which saw evolution as the spread of genetic variants that coded for favorable traits.
The tendency of an organism to develop new traits as a reaction to new environmental circumstances.
The tendency of an organism to stay the same even when exposed to new environmental circumstances.
Waddington also recognized that cells had two types of inheritance patterns. Through mitosis, one cell becomes two cells that contain the same genetic information as one another and as the original cell. The faithful transmission of the DNA base sequences is genetic transmission. And yet, genetically identical nerve cells, skin cells, and white blood cells faithfully transmit their identities as nerve cells, skin cells, and white blood cells to their descendant cells, in spite of being genetically identical (see Figure 2.14). White blood cells only make more white blood cells, never nerve cells—even though they have exactly the same DNA sequence. Waddington called this kind of cellular inheritance epigenetic.
The study of how genetically identical cells and organisms (with the same DNA base sequence) can nevertheless differ in stably inherited ways.
Here are five kinds of cells that all have the same DNA sequence yet look different.
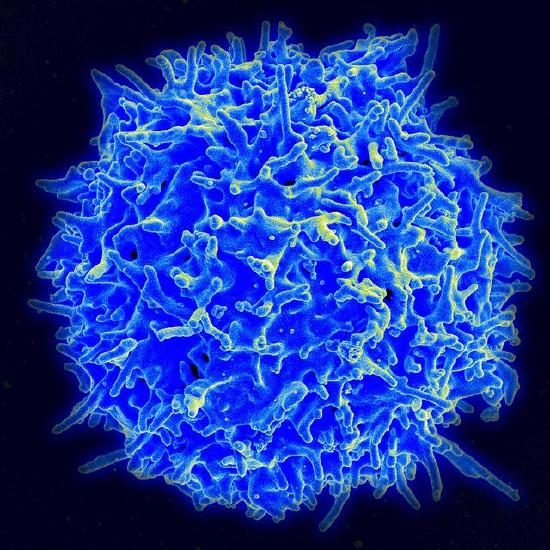
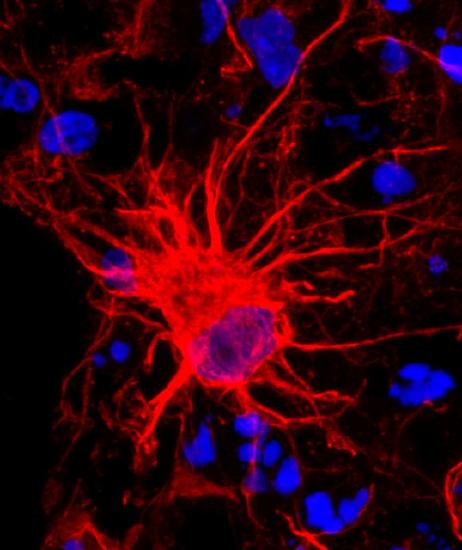
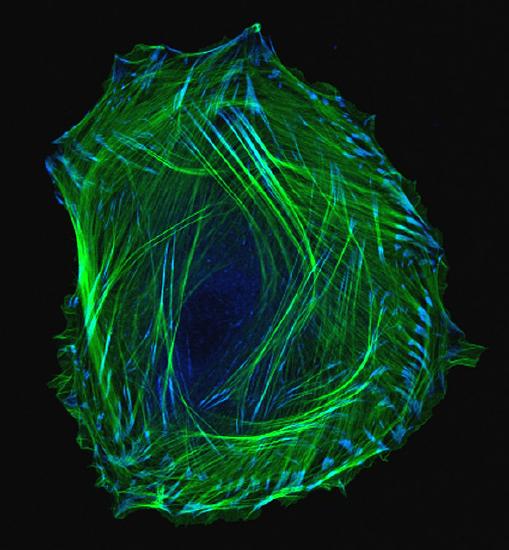
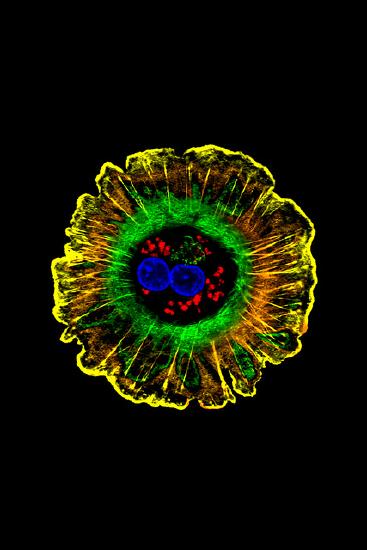
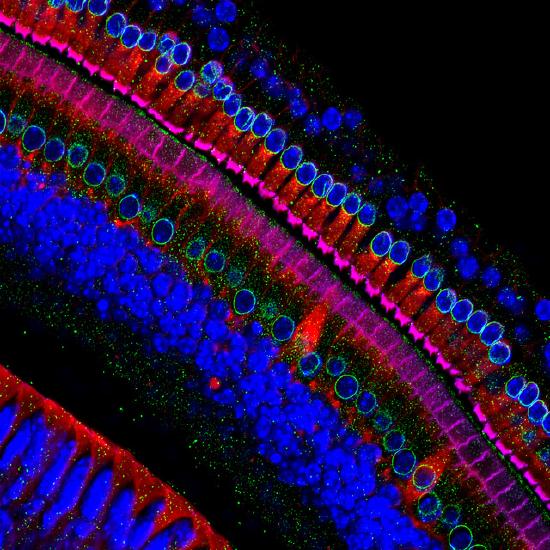
The Human Genome Project in the 1990s generated a great deal of public interest in analyzing the human DNA sequence from the standpoint of medical genetics. Some of the rhetoric was extravagant in trying to sell the public on the idea of investing a lot of money and resources in sequencing the human genome: showing the genetic basis of heritable traits, curing genetic diseases, and learning what it means ultimately to be biologically human. However, the human DNA sequence was not actually able to answer those questions, and interest began to shift from genetic information to epigenetic information: the modification of DNA structure, but not the base sequence, and the stable multi-generational inheritance of that modification.
This interest in genetics built upon decades of research in human biology, which saw the human body as highly adaptable, as controlled anthropometric studies of immigrant communities begun by anthropologists like Franz Boas and Harry Shapiro had been showing since the early 20th century. The growing human body adjusts itself to the conditions of life, such as diet, sunshine, high altitude, hard labor, population density, how babies are carried—any and all of which can have subtle but consistent effects upon its development. There can thus be no normal human form, only a context-specific range of human forms. What the human biologists called human adaptability, evolutionary biologists called developmental plasticity, and evidence quickly began to mount for its cause being epigenetic modifications to DNA.
Evolution is about how descendants come to differ from ancestors. Inheritance from parent to offspring is still the critical elementary process. But in the 21st century, the intimate relationship between evolution and inheritance has been broadened to include not merely genetic inheritance patterns but epigenetic inheritance patterns as well. We also recognize two other forms of intergenerational transmission and inheritance, which also have consequences for evolution. In addition to genetic and epigenetic variation as sources of heritable physical differences among organisms that can lead to biases in survival and reproduction, we can also model the effects of behavioral variation. Here the transmitted information is not in the DNA at all and is thus not transmitted across generations (intergenerationally). Instead, this information is transmitted horizontally (intragenerationally), permitting more rapid ways for organisms to adjust to the environment. Finally, humans are unique in that we are the only species that horizontally transmits an arbitrary set of rules to govern communication, social interaction, and thought. This shared information is symbolic and has resulted in what we recognize as “culture”: an world of names, words, pictures, classifications, revered pasts, possible futures, spirits, dead ancestors, unborn descendants, in-laws, politeness, taboo, justice, beauty, and story, all accompanied by a material world of tools. This is a fourth, symbolic or cultural mode of transmission.
Consequently, our post-Synthesis ideas about evolution tend to see the evolutionary processes as hierarchically organized and not restricted to simply the differential transmission of DNA sequences into the next generation. While that is indeed a significant part of evolution, the organism and species are nevertheless crucial to understanding how those DNA sequences get transmitted and cannot be taken for granted. Nor can we take for granted the complex roles played by the transmission of epigenetic, behavioral, and symbolic information in perpetuating our genes, bodies, and species. In the case of human evolution, one can readily see that symbolic information and cultural adaptation are far more central to our lives and our survival today than DNA and genetic adaptation. It is thus misleading to think of humans passively occupying an environmental niche. Rather, humans are actively engaged in constructing our own niches, as well as adapting to them and using them to adapt. The complex interplay between a species and its active engagement in creating its own ecology is known as niche construction.
The active engagement by which species transform their surroundings in favorable ways, rather than passively inhabiting them.
REFERENCE
Eldredge, N., and S. J. Gould. 1972. “Punctuated equilibria: an alternative to phyletic gradualism.” In Models in Paleobiology, ed.by T. J. Schopf, 82-115. San Francisco: W. H. Freeman.
FIGURE ATTRIBUTIONS
Figure 2.3.1a Horse (pferd-tier-säugetier-reiten-153500) by openclipart-vectors-30363 has been designated to the public domain (CC0).
Figure 2.3.1b Donkey by papapishu has been designated to the public domain (CC0).
Figure 2.3.2 Dentition of Ramapithecus – Fossil – Human Evolution Gallery – Indian Museum – Kolkata 2014-04-04 4499 by Biswarup Ganguly is under a CC BY-NC 4.0 License.
Figure 2.3.3 Human and ape DNA comparisons original to Explorations: An Open Invitation to Biological Anthropology is under a CC BY-NC 4.0 License.
Figure 2.3.4 Phyletic gradualism vs. punctuated equilibria original to Explorations: An Open Invitation to Biological Anthropology is under a CC BY-NC 4.0 License.
Figure 2.3.5 Mouse and elephant skeletons original to Explorations: An Open Invitation to Biological Anthropology is under a CC BY-NC 4.0 License.
Figure 2.3.6a Healthy human t cell by NIAID is under a CC BY 2.0 License.
Figure 2.3.6b Star shaped brain cells (astrocytes) by NIH (Pasca Lab, Stanford University) is under a CC BY-NC 2.0 License.
Figure 2.3.6c Embryonic smooth muscle cell by NIH (National Institute of Dental and Craniofacial Research, National Institutes of Health) is under a CC BY-NC 2.0 License.
Figure 2.3.6d Human liver cell by NIH (Donna Beer Stolz, University of Pittsburgh) is under a CC BY-NC 2.0 License.
Figure 2.3.6e Hair cells: the sound-sensing cells in the ear by NIH (Henning Horn, Brian Burke and Colin Stewart, Institute of Medical Biology, Agency for Science, Technology, and Research, Singapore) is under a CC BY-NC 2.0 License.