Charles Darwin explained in material, naturalistic terms how animals adapt to their environments. The fittest, it seems, have survived over eons of the history of life on earth to co-create ecosystems full of animals and plants. Our own bodies are full of evident adaptations: eyes for seeing, ears for hearing, and feet for walking on.
But what about hands? Feet are adapted to be primarily weight-bearing structures (rather than grasping structures, as in the apes) and that is what we primarily use them for. But we use our hands in many ways: for fine-scale manipulation, greeting, pointing, stimulating a sexual partner, writing, throwing, and cooking, among other uses. So which of these uses express what hands are “for,” when all of them express what hands do?
Figure \(\PageIndex{1}\): Chimpanzee hand (right) compared to a human hand (left).
There is an important lesson in recognizing that what things do in the present is not a good guide to understand why they came to exist. Gunpowder was invented for entertainment—and only later adopted for killing people. The Internet was invented to decentralize computers in case of a nuclear attack—and only later adopted for social media. The apes have short thumbs and use their hands in locomotion; our ancestors stopped using their hands in locomotion about six million years ago and had fairly modern-looking hands by about two million years ago. We can speculate that a combination of selection for abstract thought and dexterity led to the evolution of the human hand, with its capability for tool-making that exceeds what apes can do (see Figure 2.5.1). But let’s face it—how many tools have you made today?
Consequently, we are obliged to see the human foot as having a purpose to which it is adapted and the human hand as having multiple purposes, most of which are different from what it originally evolved for. Paleontologists Stephen Jay Gould and Elisabeth Vrba suggested that an original use be regarded as an adaptation, and the additional uses be called exaptations. Thus, we would consider the human hand to be an adaptation for tool-making and an exaptation for writing. So how do we know whether any particular feature is an adaptation, like the walking foot, rather than an exaptation, like the writing hand? Or more broadly, how can we reason rigorously from what a feature does to what it evolved for?
Definition: exaptations
An additional beneficial use for a biological feature.
The answer to the question “what did this feature evolve for?” is an origin myth. This origin myth contains three assumptions: (1) that features can be isolated and decontextualized as evolutionary units; (2) that there is a reason for the existence of any particular feature; and (3) that such a reason can be discerned.
Figure \(\PageIndex{2}\): According to the early 19th century science of phrenology, units of personality could be reliably mapped onto units of the head.
The first assumption was appreciated a century ago as the “unit-character problem.” Are the units by which the body grows and evolves the same as units we name? Clearly not; we have genes and we have noses, and we have genes that affect noses, but we don’t have “nose genes.” What, then, is the relationship between the evolving elements that we see, identify, and name and the elements that actually biologically exist and evolve? It is hard to know, but we can use the history of science as a guide to see how that fallacy has been used by earlier generations. Back in the 19th century, the early anatomists argued that since the brain contained the mind, they could map different mental states (acquisitiveness, punctuality, sensitivity) on to parts of the brain. Someone who was very introspective, say, would have an enlarged introspection part of the brain, a cranial bulge to represent the hyperactivity of this mental state. The anatomical science was known as phrenology, and it was predicated on the false assumption that units of thought or personality or behavior could be mapped to distinct parts of the brain and physically observed (see Figure 2.5.2). This is the fallacy of reification, imagining that something named is something real.
Definition: phrenology
The 19th century anatomical study of bumps on the head as an indication of personality and mental abilities.
Figure \(\PageIndex{3}\): Chimpanzees have big ears, although we don’t know why.
The second assumption, that everything has a reason, has long been recognized as a core belief of religion. Our desire to impose order and simplicity on the workings of the universe, however, does not constrain it to obey simple and orderly causes. Magic, witchcraft, spirits, and divine agency are all powerful explanations for why things happen. Consequently, it is probably not a good idea to lump natural selection in with those. Sometimes things do happen for a reason, of course, but other times things happen as byproducts of other things, or for very complicated and entangled reasons, or for no reason at all. What phenomena have reasons and thereby merit explanation? Chimpanzees have very large testicles, and we think we know why: their promiscuous sexual behavior triggers intense competition for high sperm count. But chimpanzees also have very large ears, and we don’t even try to explain them (see Figure 2.5.3). Why not? Why should there be a reason for chimp testicles but not for chimp ears? What determines the kinds of features that we try to explain, as opposed to the ones that we do not? Again, the assumption that any specific feature has a reason is metaphysical; that is to say, it may be true in any particular case, but to assume it in all cases is gratuitous.
And third, the possibility of knowing what the reason for any particular feature is, assuming that it has one, is a challenge for evolutionary epistemology (the theory of how we know things). Consider the big adaptations of our lineage: bipedalism and language. Nobody doubts that they are good and they evolved by natural selection, and we know how they work. But why did they evolve? If talking and walking are simply better than not talking and not walking, then why did they evolve in just a single branch of the ape lineage in the primate family tree? We don’t know what bipedalism evolved for, although there are plenty of speculations: walking long distances, running long distances, cooling the head, seeing over tall grass, carrying babies, carrying food, wading, threatening, counting calories, sexual display. Neither do we know what language evolved for, although there are speculations: coordinating hunting, gossiping, manipulating others. But it is also possible that bipedality is simply the way that a small arboreal ape travels on the ground, if it isn’t in the treetops. Or that language is simply the way that a primate with small canine teeth and certain mental propensities comes to communicate. If that were true, then there might be no reason for bipedality or language: having the unique suite of preconditions and a fortuitous set of circumstances simply set them in motion, and natural selection elaborated and explored their potentials. Possibly, walking and talking solved problems that no other lineage had ever solved; but even if so, the fact remains that rest of the species in the history of life have done pretty well without having solved them.
It is certainly very optimistic to think that all three assumptions (that organisms can be meaningfully atomized, that everything has a reason, and that we can know the reason) would be simultaneously in effect. Indeed, just as there are many ways of adapting (genetically, epigenetically, behaviorally, culturally), there are also many ways of being nonadaptive, which would imply that there is no reason at all for the feature in question.
First, there is the element of randomness of population histories. There are more cases of sickle-cell anemia among sub-Saharan Africans than other peoples, and there is a reason for it: carriers of sickle-cell anemia have a resistance to malaria, which is more frequent in parts of Africa (as discussed in Chapters 4 and 14). But there are more cases of a blood disease called variegated porphyria, a rare genetic metabolic disorder, in the Afrikaners of South Africa (descendants of mostly Dutch settlers in the 17th century) than in other peoples, and there is no reason for it. Yet we know the cause: One of the founding Dutch colonial settlers had the allele, and everyone in South Africa with it today is her descendant. But that is not a reason, that is simply an accident of history.
Second, there is the potential mismatch between the past and the present. The value of a particular feature in the past may be changed as the environmental circumstances change. Our species is diurnal, and our ancestors were diurnal. But beginning around a few hundred thousand years ago, our ancestors could build fires, which extended the light period, which was subsequently further amplified by lamps and candles. And over the course of the 20th century, electrical power has made it possible for people to stay up very late when it is dark—working, partying, worrying—to a greater extent than any other closely related species. In other words, we evolved to be diurnal, yet we are now far more nocturnal than any of our recent ancestors or close relatives. Are we adapting to nocturnality? If so, why? Does it even make any sense to speak of the human occupation of a nocturnal ape niche, despite the fact that we empirically seem to be doing just that? And if so, does it make sense to ask what the reason for it is?
Third, there is a genetic phenomenon known as a selective sweep, or the hitchhiker effect. Imagine three genes—A, B, and C—located very closely together on a chromosome. They each have several variants, or alleles, in the population. Now, for whatever reason, it becomes beneficial to have one of the B alleles, say B4; this B4 allele is now under strong positive selection. Obviously, we will expect future generations to be characterized by mostly B4. But what was B4 attached to? Because whatever A and C alleles were adjacent to it will also be quickly spread, simply by virtue of the selection for B4. Even if the A and C alleles are not very good, they will spread because of the good B4 allele between them. Eventually the linkage groups will break up because of genetic crossing-over in future generations. But in the meantime, some random version of genes A and C are proliferating in the species simply because they are joined to superior allele B4. And clearly, the A and C alleles are there because of selection—but not because of selection for them!
Figure \(\PageIndex{4}\): Lower jaw of Gigantopithecus.
Fourth, why does the jaw of the Miocene ape Gigantopithecus contain a first molar the size of a quarter? Was there something special about the enlarged molar? No, it had enormous jaws and teeth, and the first molar is simply one of them. This is the correlation of parts, the problem with atomizing the organism and imagining the parts to be existing and evolving independently. There is no reason for Gigantopithecus to have a large molar; there may well have been a reason that Gigantopithecus’s jaws (and, inferentially, head and body) were huge, but framing questions about the size of one tooth will never produce the correct answer (see Figure 2.19).
Fifth, some features are simply consequences of other properties rather than adaptations to external conditions. We already have noted the phenomenon of allometric growth, in which some physical features have to outgrow others simply to maintain function at an increased size. Can we ask the reason for the massive brow ridges of Homo erectus, or are brow ridges simply what you get when you have a conjunction of thick skull bones, a large face, and a sloping forehead—and, thus, again would have a cause but no reason?
Sixth, some features may be underutilized and on the way out. What is the reason for our two outer toes? They aren’t propulsive, they don’t do anything, and sometimes they’re just in the way. Obviously they are there because we are descended from pentadactyl tetrapod ancestors. Is it possible that a million years from now, we will just have our three largest toes, just as the ancestors of the horse lost their digits in favor of a single hoof per limb? Or will our outer toes find another use, such as stabilizing the landings in our personal jet-packs? For the time being, we can just recognize vestigiality as another nonadaptive explanation for the presence of a given feature.
Finally, Darwin himself recognized that many obvious features do not help an animal survive. Some things may instead help an animal breed. The peacock’s tail feathers do not help it eat, but they do help it mate. There is competition, but only against half of the species; Darwin called this sexual selection. Its result is not a fit to the environment but, rather, a fit to the opposite sex. In some species, that is literally the case, as the male and female genitalia have specific ways of anatomically fitting together. The specific form is less important than the specific match, so inquiring about the reason for a particular form of the reproductive anatomy may be misleading. The specific form may be effectively random, as long as it fits the opposite sex and is different from the anatomies of other species. Nor is sexual selection the only form of selection that can affect the body differently from natural selection. Competition might also take place between biological units other than organisms—perhaps genes, perhaps cells, or populations, or species. The spread of cultural things, such as head-binding or cheap refined fructose or forced labor, can have significant effects upon bodies, which are also not adaptations produced by natural selection. They are often adaptive physiological responses to stresses but not the products of natural selection.
Definition: sexual selection
Natural selection arising through preference by one sex for certain characteristics in individuals of the other sex.
Clearly, with so many paths available by which a physical feature might have naturalistically arisen without specifically having been the object of natural selection, it is unwise to simply assume that any individual trait is an adaptation. And that generalization applies to the best-known, best-studied, and most materially based evolutionary adaptations of our lineage. But our cultural behaviors are also highly adaptive, so what about our most familiar social behaviors? Patriarchy, hierarchy, warfare—are these adaptations? Do they have reasons? Are they good for something?
This is where some sloppy thinking has been troublesome. What would it mean to say that patriarchy evolved by natural selection in the human species? If, on the one hand, it means that the human mind evolved by natural selection to be able to create and survive in many different kinds of social and political regimes, of which patriarchy is one (or several), then biological anthropologists will readily agree. If, on the other hand, it means that patriarchy itself evolved by natural selection, that implies that patriarchy is genetically determined (since natural selection is a genetic process) and out-reproduced the alleles for other, more egalitarian, social forms. This in turn would imply that patriarchy is an adaptation and therefore of some beneficial value in the past as well as an ingrained part of human nature today. This would be bad news, say, if you harbored ambitions of dismantling it. Dismantling patriarchy in that case would be to go against nature, a futile gesture. In other words, this latter interpretation would be a naturalistic manifesto for a conservative political platform: don’t try to dismantle the patriarchy, because it is within us, the product of evolution—suck it up and live with it.
Here, evolution is being used simply as a political instrument for transforming the human genome into an imaginary glass ceiling against equality. There is thus a convergence between the pseudo-biology of crude adaptationism (the idea that everything is the product of natural selection) and the pseudo-biology of hereditarianism. Naturalizing inequality is not the business of evolutionary theory, and it represents a difficult moral position for a scientist to adopt, as well as a poor scientific position.
Definition: adaptationism
The idea that everything is the product of natural selection.
FIGURE ATTRIBUTIONS
Figure 2.5.1 Human and chimpanzee hand original to Explorations: An Open Invitation to Biological Anthropology is under a CC BY-NC 4.0 License.
Figure 2.5.2 Phrenology; Chart (slide number 5278, photo number: L0000992) by Wellcome Collection, original from The Phrenological Journal (Know Thyself), print from Dr. E. Clark is under a CC BY 4.0 license.

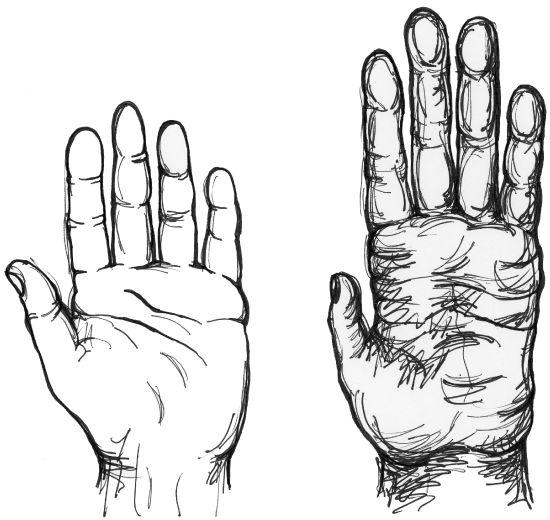 Figure \(\PageIndex{1}\): Chimpanzee hand (right) compared to a human hand (left).
Figure \(\PageIndex{1}\): Chimpanzee hand (right) compared to a human hand (left).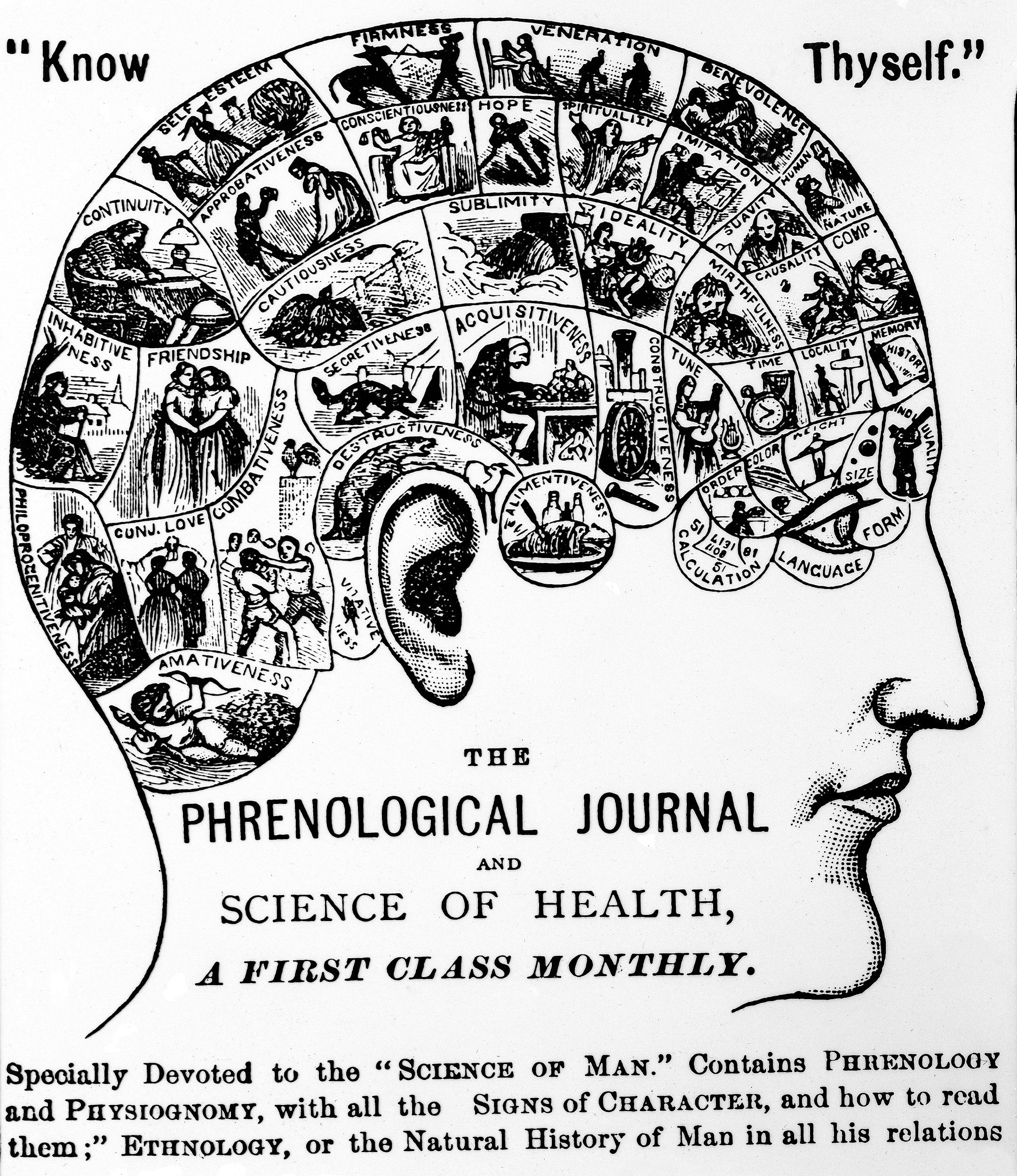 Figure \(\PageIndex{2}\): According to the early 19th century science of phrenology, units of personality could be reliably mapped onto units of the head.
Figure \(\PageIndex{2}\): According to the early 19th century science of phrenology, units of personality could be reliably mapped onto units of the head. Figure \(\PageIndex{3}\): Chimpanzees have big ears, although we don’t know why.
Figure \(\PageIndex{3}\): Chimpanzees have big ears, although we don’t know why.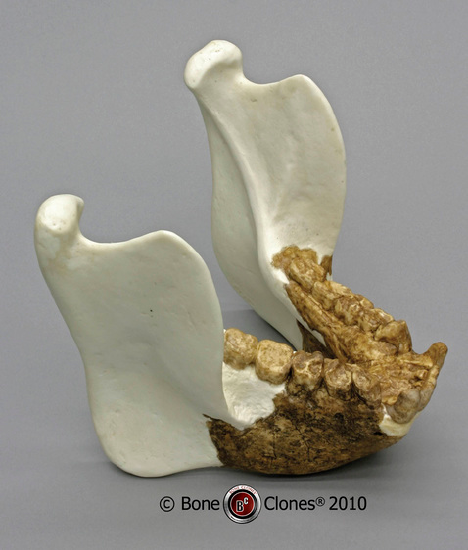 Figure \(\PageIndex{4}\): Lower jaw of Gigantopithecus.
Figure \(\PageIndex{4}\): Lower jaw of Gigantopithecus.