
8.1: What Is A Primate?
- Page ID
- 158763
MORE ON PRIMATES
The Order: Plesiadapiformes. Archaic primates or primate-like placental mammals (Early Paleocene–Late Eocene).
The Order: Primates. True primates or primates of modern aspect.
Group containing monkeys and apes, including humans.
The continents of Africa and Eurasia.
How to Diagnose a Primate
The identifying features that allow you to recognize a member of a group.
The Order: Primates; Suborder: Anthropoidea; Infraorder: Catarrhini. The group containing Old World monkeys and apes, including humans.
The Order: Primates; Suborder: Anthropoidea; Infraorder: Platyrrhini. The group containing New World monkeys.
MAJOR HYPOTHESES ABOUT PRIMATE ORIGINS
For many groups of mammals, there is a key feature that led to their success. A good example is powered flight in bats. Primates lack a feature like this. Instead, if there is something unique about primates, it is probably a group of features rather than one single thing. Because of this, anthropologists and paleontologists struggle to describe an ecological scenario that could explain the rise and success of our own order. Three major hypotheses have been advanced to explain the origin of primates and to explain what makes our own order unique among mammals (Figure 8.1.1); these are described below.
 Figure \(\PageIndex{1}\): Three major hypotheses are A) the arboreal hypothesis, B) the visual predation hypothesis, and C) the angiosperm-primate coevolution hypothesis.
Figure \(\PageIndex{1}\): Three major hypotheses are A) the arboreal hypothesis, B) the visual predation hypothesis, and C) the angiosperm-primate coevolution hypothesis.Arboreal Hypothesis
In the 1800s, many anthropologists viewed all animals in relation to humans. That is, animals that were more like humans were considered to be more “advanced” and those lacking humanlike features were considered more “primitive.” This way of thinking was particularly obvious in studies of primates. Thus, when anthropologists sought features that separate primates from other mammals, they focused on features that were least developed in lemurs and lorises, more developed in monkeys, and most developed in apes (Figure 8.1.2). Frederic Wood Jones, one of the leading anatomist-anthropologists of the early 1900s, is usually credited with the Arboreal Hypothesis of primate origins (Jones 1916). This hypothesis holds that many of the features of primates evolved to improve locomotion in the trees. For example, the grasping hands and feet of primates are well suited to gripping tree branches of various sizes and our flexible joints are good for reorienting the extremities in many different ways. A mentor of Jones, Grafton Elliot Smith, had suggested that the reduced olfactory system, acute vision, and forward-facing eyes of primates are an adaptation to making accurate leaps and bounds through a complex, three-dimensional canopy (Smith 1912). The forward orientation of the eyes in primates causes the visual fields to overlap, enhancing depth perception, especially at close range. Evidence to support this hypothesis includes the facts that many extant primates are arboreal, and the primitive members of most primate groups are dedicated arborealists. The Arboreal Hypothesis was well accepted by most anthropologists at the time and for decades afterward.
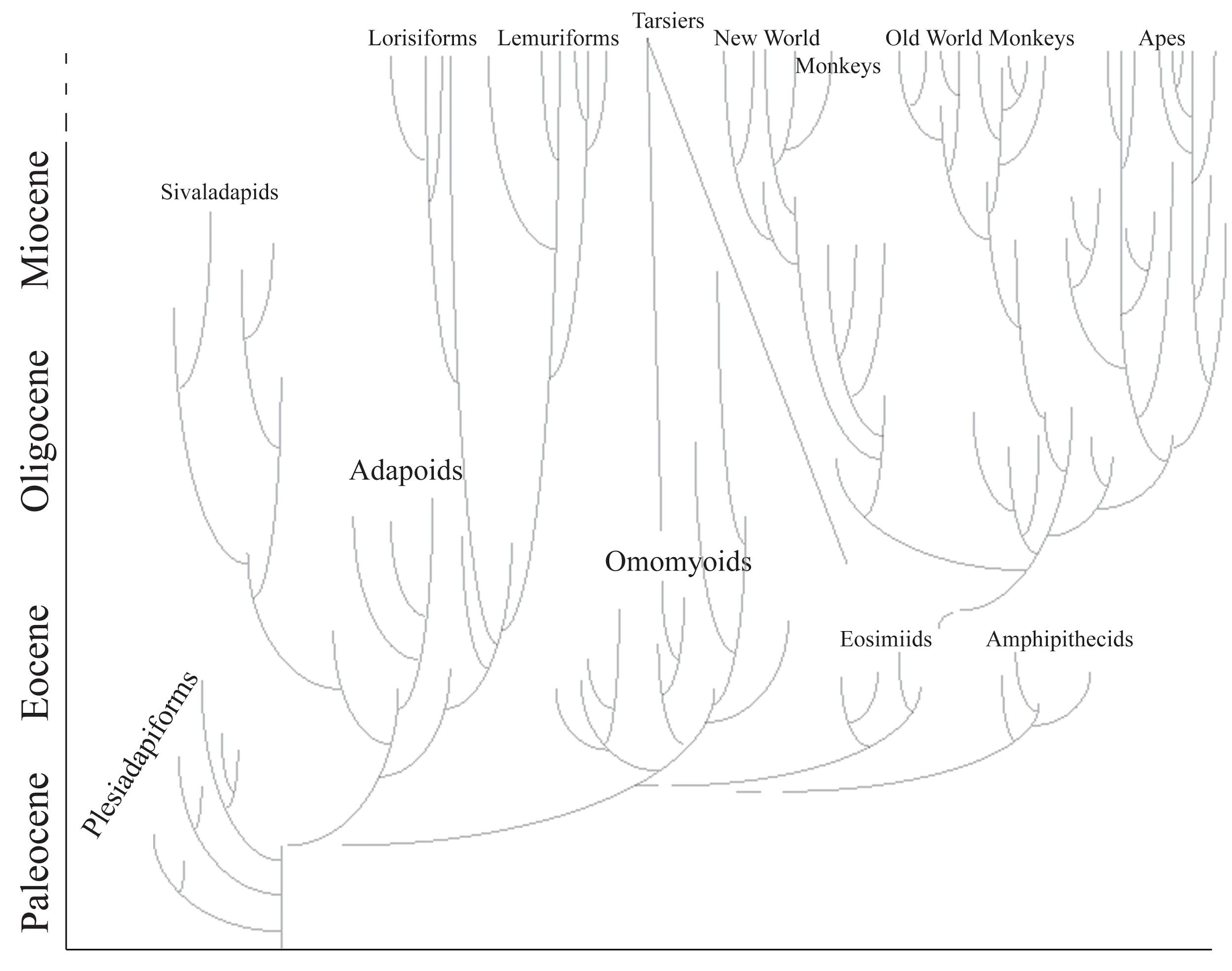 Figure \(\PageIndex{2}\): Primate family tree showing major groups. Disconnected lines show uncertainty about relationships. Note two lines leading to tarsiers from different possible groups of origin. The timescale is shortened for the epochs since the Miocene.
Figure \(\PageIndex{2}\): Primate family tree showing major groups. Disconnected lines show uncertainty about relationships. Note two lines leading to tarsiers from different possible groups of origin. The timescale is shortened for the epochs since the Miocene.Visual Predation Hypothesis
In the late 1960s and early 1970s, Matt Cartmill studied and tested the idea that the characteristic features of primates evolved in the context of arboreal locomotion. Cartmill noted that squirrels climb trees (and even vertical walls) very effectively, even though they lack some of the key adaptations of primates. As members of the Order Rodentia, squirrels also lack the hand and foot anatomy of primates. They have claws instead of flattened nails and their eyes face more laterally than those of primates. Cartmill reasoned that there must be some other explanation for the unique traits of primates. He noted that some non-arboreal animals share at least some of these traits with primates; for example, cats and predatory birds have forward-facing eyes that enable visual field overlap. Cartmill suggested that the unique suite of features in primates is an adaptation to detecting insect prey and guiding the hands (or feet) to catch insects (Cartmill 1972). His hypothesis emphasizes the primary role of vision in prey detection and capture; it is explicitly comparative, relying on form function relationships in other mammals and nonmammalian vertebrates. According to Cartmill, many of the key features of primates evolved for preying on insects in this special manner (Cartmill 1974).
Angiosperm-Primate Coevolution Hypothesis
The visual predation hypothesis was unpopular with some anthropologists. One reason for this is that many primates today are not especially predatory. Another is that, whereas primates do seem well adapted to moving around in the smallest, terminal branches of trees, insects are not necessarily easier to find there. A counterargument to the visual predation hypothesis is the angiosperm-primate coevolution hypothesis. Primate ecologist Robert Sussman (Sussman 1991) argued that the few primates that eat mostly insects often catch their prey on the ground rather than in the fine branches of trees. Furthermore, predatory primates often use their ears more than their eyes to detect prey. Finally, most early primate fossils show signs of having been omnivorous rather than insectivorous. Instead, he argued, the earliest primates were probably seeking fruit. Fruit (and flowers) of angiosperms (flowering plants) often develop in the terminal branches. Therefore, any mammal trying to access those fruits must possess anatomical traits that allow them to maintain their hold on thin branches and avoid falling while reaching for the fruits. Primates likely evolved their distinctive visual traits and extremities in the Paleocene (approximately 65 million to 54 million years ago) and Eocene (approximately 54 million to 34 million years ago) epochs, just when angiosperms were going through a revolution of their own—the evolution of large, fleshy fruit that would have been attractive to a small arboreal mammal. Sussman argued that, just as primates were evolving anatomical traits that made them more efficient fruit foragers, angiosperms were also evolving fruit that would be more attractive to primates to promote better seed dispersal. This mutually beneficial relationship between the angiosperms and the primates was termed “coevolution” or more specifically “diffuse coevolution.”
The ecological interaction between whole groups of species (e.g., primates) with whole groups of other species (e.g., fruiting trees).
At about the same time, D. Tab Rasmussen noted several parallel traits in primates and the South American woolly opossum, Caluromys. He argued that early primates were probably foraging on both fruits and insects (Rasmussen 1990). As is true of Caluromys today, early primates probably foraged for fruits in the terminal branches of angiosperms, and they probably used their visual sense to aid in catching insects. Insects are also attracted to fruit (and flowers), so these insects represent a convenient opportunity for a primarily fruit-eating primate to gather protein. This solution is, in effect, a compromise between the visual predation hypothesis and the angiosperm-primate coevolution hypothesis. It is worth noting that other models of primate origins have been proposed, and these include the possibility that no single ecological scenario can account for the origin of primates.
REFERENCES
Cartmill, Matt. 1972. Arboreal Adaptations and the Origin of the Order Primates. In The Functional and Evolutionary Biology of Primates, edited by Russell Tuttle, 97–122. Chicago: Aldine-Atherton.
Cartmill, Matt. 1974. Rethinking Primate Origins. Science 184 (4,135): 436–443.
Jones, F. Wood. 1916. Arboreal Man. London: Edward Arnold.
Martin, R. D. 1968. Towards a New Definition of Primates. Man (N.S.) 3 (3): 377–401.
Martin, R. D. 1990. Primate Origins and Evolution, a Phylogenetic Reconstruction. Princeton: Princeton University Press.
Rasmussen, D. Tab. 1990. Primate Origins: Lessons From a Neotropical Marsupial. American Journal of Primatology 22 (4): 263–277.
Smith, G. Elliot. 1912. The Evolution of Man. Smithsonian Institute Annual Report 2012: 553–572.
Sussman, Robert W. 1991. Primate Origins and the Evolution of Angiosperms. American Journal of Primatology 23 (4): 209–223.
FIGURE ATTRIBUTIONS
Figure 8.1.1 Hypotheses about primate origins a derivative work by Jonathan M. G. Perry and Stephanie L. Canington is under a CC BY-NC 4.0 License. [Includes Garnett’s Galago by Ltshears, CC BY-SA 3.0; Nycticebus bancanus by Gunay M.E., CC BY-SA 4.0; Anja réserve (Madagascar) – 06 by Wayne77, CC BY-SA 4.0].
Figure 8.1.2 Primate family tree by Jonathan M. G. Perry is under a CC BY-NC 4.0 License.