
9.2: Derived Adaptations
- Page ID
- 158772
BIPEDALISM
Where the primary form of locomotion for an organism is bipedal.
Maintaining body temperature through physiologically cooling or warming the body.
Skeletal Adaptations for Bipedalism
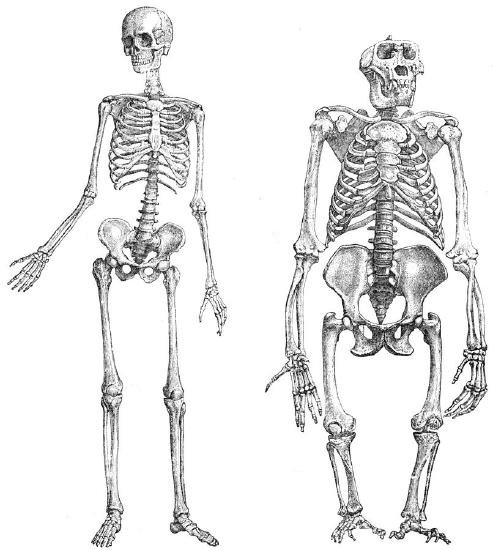 Figure \(\PageIndex{1}\): Compared to gorillas (right) and other apes, humans (left) have highly specialized adaptations to facilitate bipedal locomotion.
Figure \(\PageIndex{1}\): Compared to gorillas (right) and other apes, humans (left) have highly specialized adaptations to facilitate bipedal locomotion.The skeleton below the cranium (head).
| Region | Feature | Obligate Biped (H. sapiens) | Non-obligate Biped |
| Cranium | Position of the foramen magnum | Positioned inferiorly (immediately under the cranium) so that the head rests on top of the vertebral column for balance and support (head is perpendicular to the ground | Posteriorly positioned (to the back of the cranium). Head is positioned parallel to the ground |
| Postcranium | Body proportions | Shorter upper limb (not used for locomotion) | Nonhuman apes: Longer upper limbs (used for locomotion) |
| Postcranium | Spinal curvature | S-curve due to pressure exerted on the spine from bipedalism (lumbar lordosis) | C-curve |
| Postcranium | Vertebrae | Robust lumbar (lower-back) vertebrae (for shock absorbance and weight bearing). Lower back is more flexible than that of apes as the hips and trunk swivel when walking (weight transmission). | Gracile lumbar vertebrae compared to those of modern humans |
| Postcranium | Pelvis | Shorter, broader bowl-shaped pelvis (for support); very robust. Broad sacrum with large sacroiliac joint surfaces | Longer, flatter, elongated ilia, more narrow and gracile, narrower sacrum, relatively smaller sacroiliac joint surface |
| Postcranium | Lower limb | In general, longer, more robust lower limbs and more stable, larger joints
|
In general, smaller, more gracile limbs with more flexible joints
|
| Postcranium | Foot | Rigid, robust foot, without a midtarsal break. Non-opposable and large, robust big toe (for push off while walking) and large heel for shock absorbance. | Flexible foot, midtarsal break present (which allows primates to lift their heels independently from their feet), opposable big toe for grasping. |
The large hole (foramen) at the base of the cranium, through which the spinal cord enters the skull.
The inward curving of the lower (lumbar) parts of the spine. The lower curve in the human S-shaped spine.
The angle of the knee between the femur and tibia, which allows for weight distribution to be angled closer to the point above the center of gravity (i.e., between the feet) in bipeds.
The concept that different characteristics evolve at different rates and appear at different stages.
The big toe.
Long bones in the hand and fingers.
Earliest Hominins: Sahelanthropus and Orrorin
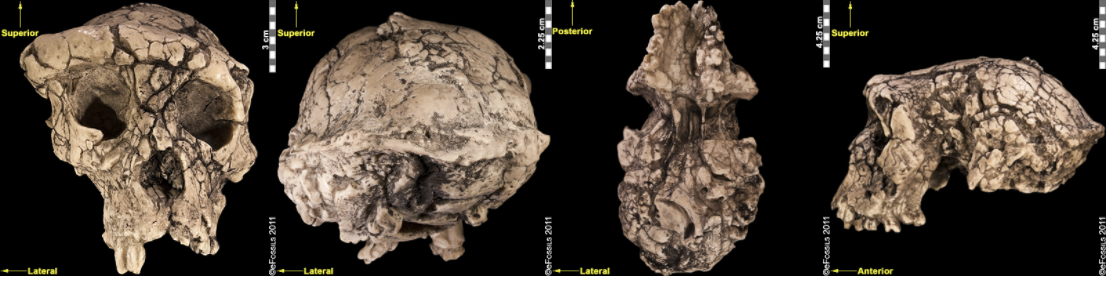 Figure \(\PageIndex{2}\): Sahelanthropus tchadensis exhibits a set of derived features, including a long, low cranium; a small, ape-sized braincase; and relatively reduced prognathism.
Figure \(\PageIndex{2}\): Sahelanthropus tchadensis exhibits a set of derived features, including a long, low cranium; a small, ape-sized braincase; and relatively reduced prognathism.The hind dentition (molars and premolars).
A geological epoch between the Miocene and Pleistocene.
Earliest Hominins: The Genus Ardipithecus
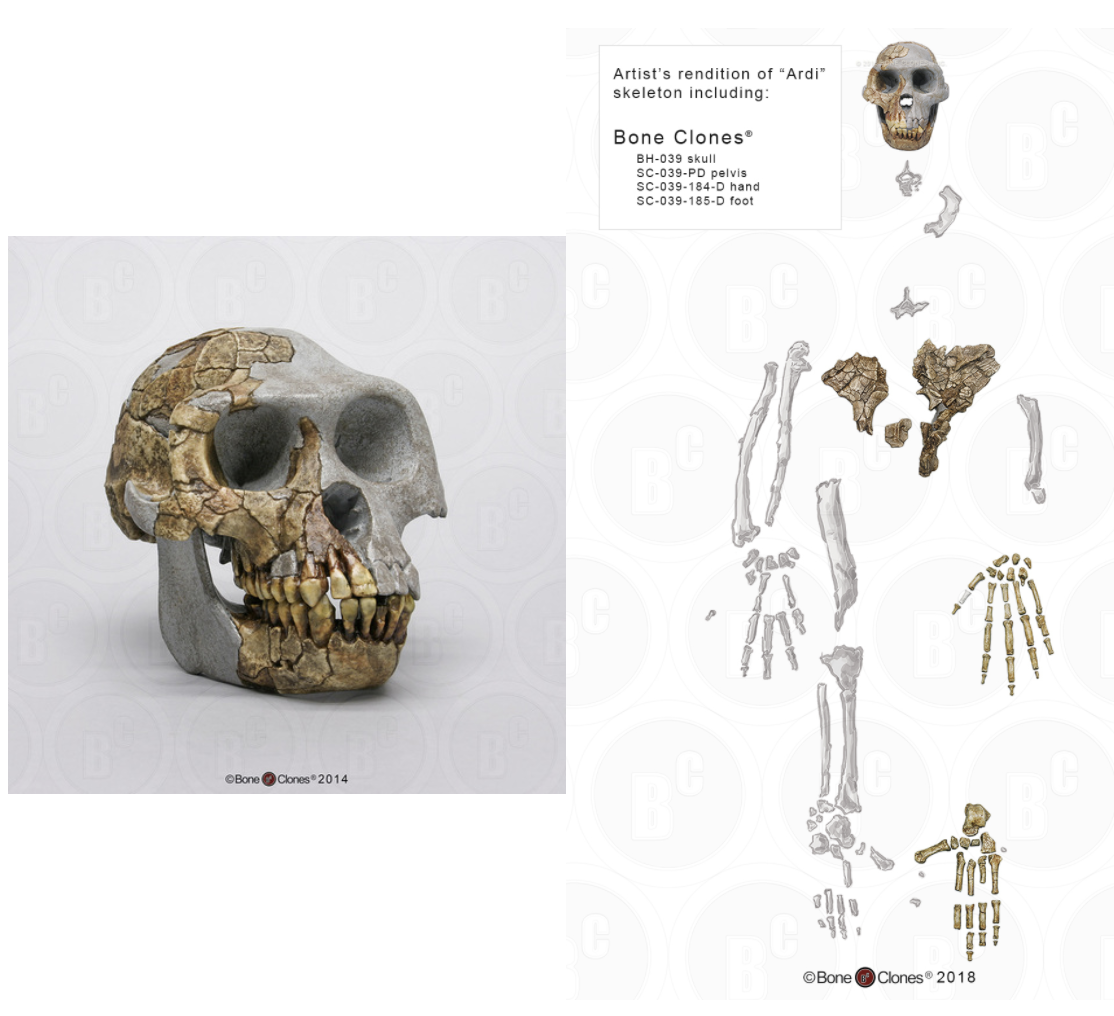 Figure \(\PageIndex{3}\): Researchers believe that Ardipithecus ramidus was able to walk upright, although not as efficiently as later humans. It possessed the musculature required for tree climbing, and while moving quadrupedally, it likely placed weight on the palms of the hands rather than on the knuckles.
Figure \(\PageIndex{3}\): Researchers believe that Ardipithecus ramidus was able to walk upright, although not as efficiently as later humans. It possessed the musculature required for tree climbing, and while moving quadrupedally, it likely placed weight on the palms of the hands rather than on the knuckles.Bipedal Trends in Early Hominins
EARLY HOMININ DENTITION
The Importance of Teeth
Teeth are abundant in the fossil record, primarily because they are already highly mineralized as they are forming, far more so than even bone. Because of this, teeth preserve readily. And, because they preserve readily, they are well-studied and better understood than many skeletal elements. Even in the sparse hominin (and primate) fossil record, teeth are, in some cases, all we have.
Teeth also reveal a lot about the individual from whom they came. We can tell what they evolved to eat, which other species they may be more closely related to, and even, to some extent, the level of sexual dimorphism, or general variability, within a given species. This is powerful information that can be contained in a single tooth. With a little more observation, the wearing patterns on a tooth can tell us about the diet of the individual in the weeks leading up to its death. Furthermore, the way in which a tooth is formed, and the timing of formation, can reveal information about changes in diet (or even mobility) over infancy and childhood, using isotopic analyses. When it comes to our earliest hominin relatives, this information is vital for understanding how they lived.
The purpose of comparing different hominin species is to better understand the functional morphology as it applies to dentition. In this, we mean that the morphology of the teeth or masticatory system (which includes jaws) can reveal something about the way in which they were used and, therefore, the kinds of foods these hominins ate. When comparing the features of hominin groups, it is worth considering modern analogues (i.e., animals with which to compare) to make more appropriate assumptions about diet. In this way, hominin dentition is often compared with that of chimpanzees, gorillas (our two closest relatives), and/or modern humans.
The most divergent group, however, is humans. Humans around the world have incredibly varied diets. Among hunter-gatherers, it can vary from a honey- and plant-rich diet, as seen in the Hadza in Tanzania, to a diet almost entirely reliant on animal fat and protein, as seen in Inuits in polar regions of the world. We are therefore considered generalists, more general than the largely frugivorous (fruit-eating) chimpanzee or the folivorous (foliage-eating) gorilla.
One way in which all humans are similar is our reliance on the processing of our food. We cut up and tear meat with tools using our hands, instead of using our front teeth (incisors and canines). We smash and grind up hard seeds, instead of crushing them with our hind teeth (molars). This means that, unlike our ape relatives, we can rely more on developing tools to navigate our complex and varied diets. Our brain, therefore, is our primary masticatory organ. Evolutionarily, partially in response to our increased reliance on our hands and brain, our teeth have reduced in size and our faces are flatter, or more orthognathic. Similarly, a reduction in teeth and a more generalist dental morphology could also indicate an increase in softer and more variable foods, such as the inclusion of more meat. These trends begin early on in our evolution. The link has been made between some of the earliest evidence for stone tool manufacture, the earliest members of our genus, and the features that we associate with these specimens.
The face below the eyes is relatively flat and does not jut out anteriorly.
General Dental Trends in Early Hominins
Several trends are visible in the dentition of early hominins. However, worth noting is that all tend to have the same dental formula. The dental formula is a method to characterize how many of the different kinds of teeth are present in the mouth. Going from the most anterior (front) of the mouth, this includes the square, flat incisors; the pointy canines; the small, flatter premolars; and the larger hind molars. In many primates, from Old World monkeys to great apes, the typical dental formula is 2:1:2:3. This means that if we divide the mouth into quadrants, each should have two incisors, one canine, two premolars and three molars. In total that is eight teeth a quadrant, for a total of 32 teeth. In humans, this number can be variable. Unlike in other apes, it is not uncommon for people to have only two molars in one or more of their quadrants. One explanation for this is that, because of our processed foods, there are fewer dietary constraints—that is, less pressure to have many teeth for additional processing. Furthermore, with smaller mouths and faces, fewer teeth may be advantageous. All early hominins have the primitive condition shared with other great apes.
The morphology of the individual teeth is where we see the most change. Among primates, large incisors are associated with food procurement or preparation (such as biting small fruits), while small incisors indicate a diet which may contain small seeds or leaves (where the preparation is primarily in the back of the mouth). Most hominins have relatively large, flat, vertically aligned incisors that occlude (touch) relatively well, forming a “bite.” This differs from, for instance, the orangutan, whose teeth stick out (i.e., are procumbent).
When the teeth from the maxilla come into contact with the teeth in the mandible.
In reference to incisors, tilting forward.
While the teeth are often sensitive, evolutionarily speaking, with diet, the canines may be misleading in that regard. We tend to associate pointy, large canines with the ripping required for meat, and the reduction (or, in some animals, the absence) of canines as indicative of more herbivorous diets. In humans, our canines are often a similar size to our incisors and are therefore considered incisiform (Figure \(\PageIndex{4}\)). However, our closest relatives all have very long, pointy canines, particularly on their upper dentition. This is true even for the gorilla, which lives almost exclusively on plants, as you have seen in previous chapters. The canines, in these instances, possibly indicate more about social structure and sexual dimorphism than diet.
An adjective referring to a canine that appears more incisor-like in morphology.
 Figure \(\PageIndex{4}\): In humans, our canines are often a similar size to our incisors.
Figure \(\PageIndex{4}\): In humans, our canines are often a similar size to our incisors.Early on in human evolution, we see a reduction in canine size. Sahelanthropus tchadensis and Orrorin tugenensis both have smaller canines than those in extant great apes, yet the canines are still larger and pointier than those in humans or more recent hominins. This implies strongly that, over evolutionary time, the need for display and dominance among males has reduced, as has our sexual dimorphism. In Ardipithecus ramidus, there is no obvious difference between male and female canine size, yet they are still slightly larger and pointier than in humans. This implies a less sexually dimorphic social structure in the earlier hominins relative to modern-day chimpanzees and gorillas.
Along with a reduction in canine size is the reduction or elimination of a canine diastema: a gap between the teeth on the mandible that allows room for elongated teeth on the maxilla to “fit” in the mouth. Absence of a diastema is an excellent indication of a reduction in canine size. In animals with large canines (such as baboons), there is also often a honing P3, where the first premolar (also known as P3 for evolutionary reasons) is triangular in shape, “sharpened” by the extended canine from the upper dentition. Evidence for this is also seen in some of the early hominins such as Ardipithecus, for whom even though the canines are much smaller and almost the same height as the incisors, they are larger than those in more recent hominins.
The mandibular premolar alongside the canine (in primates, the P3), which is angled to give space for (and sharpen) the upper canines.
The hind dentition, such as the bicuspid (two cusped) premolars or the much larger molars, are also highly indicative of a generalist diet in hominins. Among the earliest hominins, the molars are larger than we see in our genus, increasing in size to the back of the mouth and angled in such a way from the much smaller anterior dentition as to give these hominins a parabolic (V-shaped) dental arch. This is opposed to our living relatives as well as some of the earliest hominins, such as Sahelanthropus, whose molars and premolars are relatively parallel between the left and right sides of the mouth, creating a U-shape.
Shaped like a parabola (V-shaped).
Among more recent early hominins, the molars are relatively large, larger than those in the earliest hominins and far larger than those in our own genus, Homo. Large, short molars with thick enamel allow these early cousins of ours to grind away at fibrous, coarse foods, such as sedges, which require plenty of chewing. This is further evidenced in the low cusps, or ridges, on the teeth, which are ideal for chewing. In our genus, the hind dentition is far smaller than in these early hominins. Our teeth also have medium-size cusps, which allow for both efficient grinding and tearing/shearing meats.
Understanding the dental morphology has allowed researchers to extrapolate very specific behaviors of early hominins. It is worth noting that while teeth preserve well and are abundant, a slew of other morphological traits additionally provide evidence for many of these hypotheses. Yet there are some traits that are ambiguous. For instance, while there are definitely high levels of sexual dimorphism in Au. afarensis, which we will discuss in the next section, the canine teeth are reduced in size, implying that while canines may be useful indicators for sexual dimorphism, it is also worth noting other lines of evidence.
The highly mineralized outer layer of the tooth.
Dental Trends in Early Hominins
Trends among early hominins include a reduction in procumbency, reduced hind dentition (molars and premolars), a reduction in canine size (more incisiform with a lack of canine diastema and honing P3), flatter molar cusps, and thicker dental enamel. All early hominins have the primitive dental formula of 2:1:2:3. These trends are all consistent with a generalist diet, incorporating more fibrous foods.
REFERENCES
Brunet, M., A. Beauvilain, Y. Coppens, E. Heintz, A. H. Moutaye, and D. Pilbeam. 1995. “The First Australopithecine 2,500 Kilometers West of the Rift Valley (Chad).” Nature 378 (6554): 275–273.
White, T. D., G. Suwa, and B. Asfaw. 1994. “Australopithecus ramidus, a New Species of Early Hominid from Aramis, Ethiopia.” Nature 371(6495): 306–312.
FIGURE ATTRIBUTIONS
Figure 9.2.1 Skeleton of human (1) and gorilla (2), unnaturally sketched by unknown from Brehms Tierleben, Small Edition 1927 is in the public domain.
Figure 9.2.2 Derivitive of Sahelanthropus tchadensis: TM 266-01-060-1 anterior view, Sahelanthropus tchadensis: TM 266-01-060-1 posterior view, Sahelanthropus tchadensis: TM 266-01-060-1 inferior view, and Sahelanthropus tchadensis: TM 266-01-060-1 lateral left view by eFossils is copyrighted and used for noncommercial purposes as outlined by eFossils.
Figure 9.2.3 Derivitive of Ardipithecus ramidus Skull and Artist’s rendition of “Ardi” skeleton by ©BoneClones is used by permission and available here under a CC BY-NC 4.0 License.
Figure 9.2.4 Adult human teeth by Genusfotografen (Tomas Gunnarsson) through Wikimedia Sverige Wikimedia Sverige is used under a CC BY-SA 4.0 License.
TABLE ATTRIBUTION
Table 9.2.1 Skeletal comparisons between modern humans and non-obligate bipeds original to Explorations: An Open Invitation to Biological Anthropology is under a CC BY-NC 4.0 License.