
9.3: Australopithecus And Tool Use
- Page ID
- 158774
THE GENUS AUSTRALOPITHECUS
An organism with extremely large dentition compared with body size.
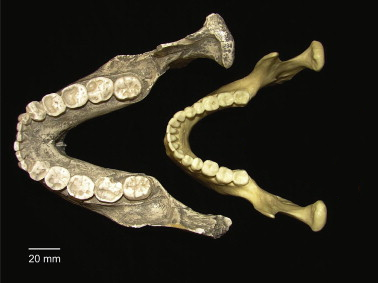 Figure \(\PageIndex{1}\): Robust Australopithecines (left) had large molars and chewing muscles compared to modern humans (right).
Figure \(\PageIndex{1}\): Robust Australopithecines (left) had large molars and chewing muscles compared to modern humans (right).Gracile Australopithecines
Slender, less rugged, or pronounced features.
Rugged or exaggerated features.
East African Australopithecines
Rock made from ash from volcanic eruptions in the past.
As pertaining to palaeoanthropology, this term refers to the place where an artifact or fossil is found.
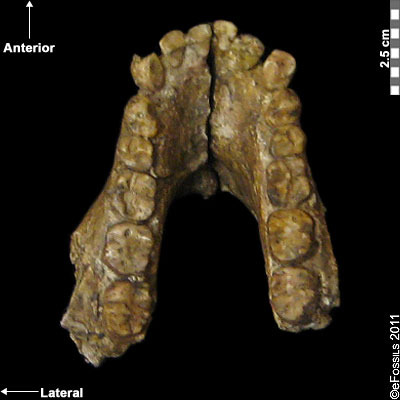 Figure \(\PageIndex{2}\): As seen in this mandible, Australopithecus anamensis had relatively large canine teeth.
Figure \(\PageIndex{2}\): As seen in this mandible, Australopithecus anamensis had relatively large canine teeth. Figure \(\PageIndex{3}\): Artistic reconstructions of Australopithecus afarensis by artist John Gurche. Female “Lucy” is left and a male is on the right.
Figure \(\PageIndex{3}\): Artistic reconstructions of Australopithecus afarensis by artist John Gurche. Female “Lucy” is left and a male is on the right.In reference to the face, the area below the eyes juts anteriorly.
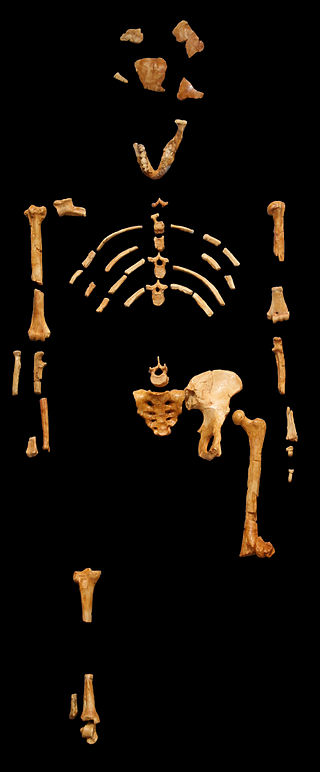 Figure \(\PageIndex{4}\): The humanlike femoral neck, valgus knee, and bowl-shaped hip seen in the “Lucy” skeleton indicates that Australopithecus afarensis was bipedal.
Figure \(\PageIndex{4}\): The humanlike femoral neck, valgus knee, and bowl-shaped hip seen in the “Lucy” skeleton indicates that Australopithecus afarensis was bipedal.Contested Species
A single specimen from which a species or taxon is described or named.
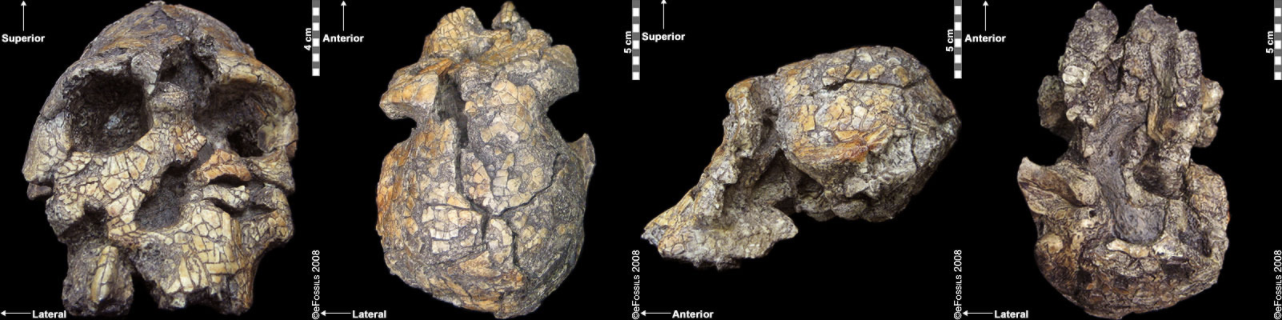 Figure \(\PageIndex{5}\): This specimen (Kenyanthopus platyops) has small detention, a small brain case, and a relatively flat face. Its genus/species designation remains contested.
Figure \(\PageIndex{5}\): This specimen (Kenyanthopus platyops) has small detention, a small brain case, and a relatively flat face. Its genus/species designation remains contested.South African Australopithecines
Hard, calcareous sedimentary rock.
 Figure \(\PageIndex{6}\): The Taung Child has a nearly complete face, mandible, and partial endocranial cast.
Figure \(\PageIndex{6}\): The Taung Child has a nearly complete face, mandible, and partial endocranial cast. Figure \(\PageIndex{7}\): An artistic reconstruction of Australopithecus africanus by John Gurche.
Figure \(\PageIndex{7}\): An artistic reconstruction of Australopithecus africanus by John Gurche.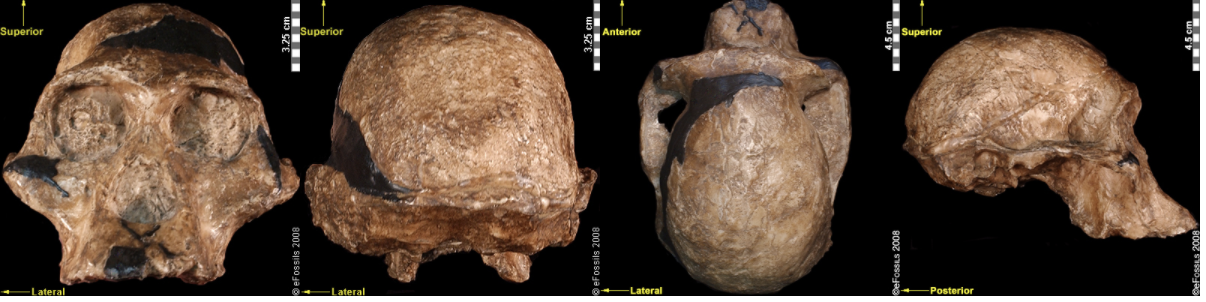 Figure \(\PageIndex{8}\): The “Mrs. Ples” brain case is small in size (like apes) but has a less prognathic face and its foramen magnum is positioned more like modern humans than in African apes.
Figure \(\PageIndex{8}\): The “Mrs. Ples” brain case is small in size (like apes) but has a less prognathic face and its foramen magnum is positioned more like modern humans than in African apes.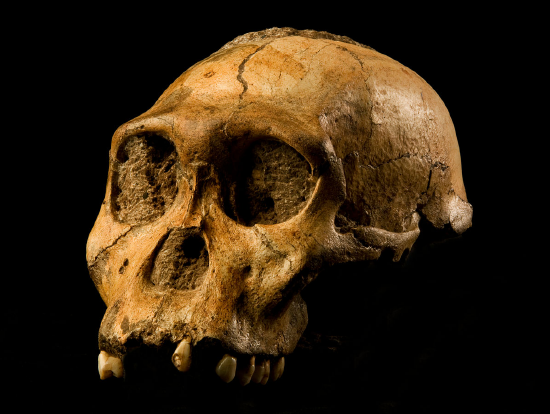 Figure \(\PageIndex{9}\): Australopithecus sediba shows mosaic features between Au. africanus and Homo.
Figure \(\PageIndex{9}\): Australopithecus sediba shows mosaic features between Au. africanus and Homo.Paranthropus “Robust” Australopithecines
Foods that may not be preferred by an animal (e.g., foods that are not nutritionally dense) but that are essential for survival in times of stress or scarcity.
A taxon or group of taxa descended from a common ancestor that is not shared with another taxon or group.
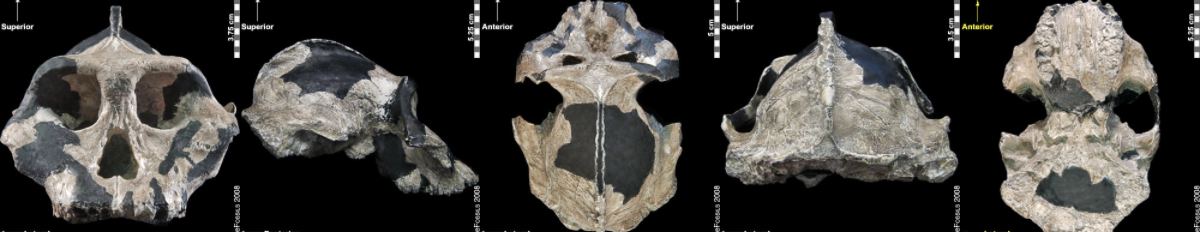 Figure \(\PageIndex{10}\): The “Black Skull” (Paranthropus aethiopicus) had a large sagittal crest and large, flared zygomatic arches that indicate it had large chewing muscles and a powerful biting force.
Figure \(\PageIndex{10}\): The “Black Skull” (Paranthropus aethiopicus) had a large sagittal crest and large, flared zygomatic arches that indicate it had large chewing muscles and a powerful biting force. Figure \(\PageIndex{11}\): Artistic reconstruction of a Paranthropus boisei, male, by John Gurche.
Figure \(\PageIndex{11}\): Artistic reconstruction of a Paranthropus boisei, male, by John Gurche.A sample (here, fossil) from which researchers extrapolate features of a population.
Even more robust than considered normal in the Paranthropus genus.
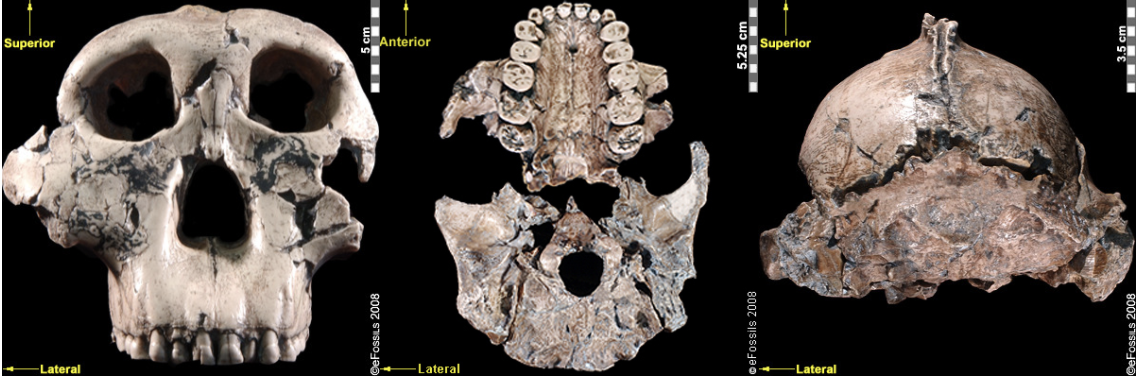 Figure \(\PageIndex{12}\): “Nutcracker Man” (Paranthropus boisei) had hyper-robust features including very large dentition, flaring zygomatic arches, a broad concave face. It had a powerful and extremely efficient chewing force.
Figure \(\PageIndex{12}\): “Nutcracker Man” (Paranthropus boisei) had hyper-robust features including very large dentition, flaring zygomatic arches, a broad concave face. It had a powerful and extremely efficient chewing force.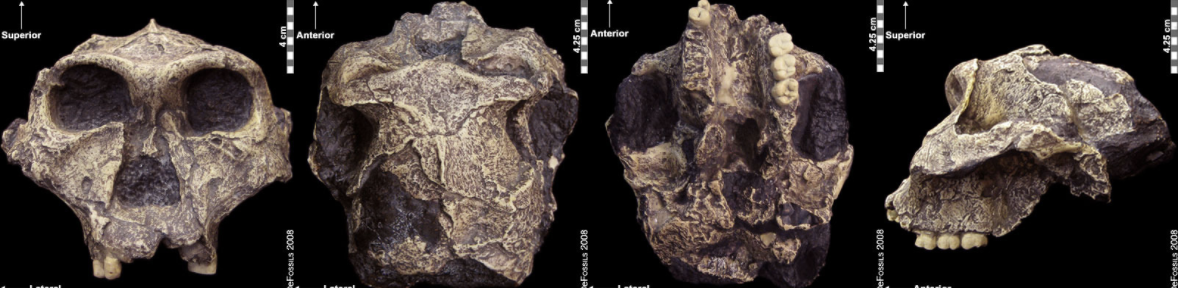 Figure \(\PageIndex{13}\): Paranthropus robustus had less derived, more general features; it was not as robust as P. boisei and not as primitive as P. aethiopicus. It also appeared to be prone to a tooth defect (enamel hypoplasia), making the enamel thinner and more prone to decay.
Figure \(\PageIndex{13}\): Paranthropus robustus had less derived, more general features; it was not as robust as P. boisei and not as primitive as P. aethiopicus. It also appeared to be prone to a tooth defect (enamel hypoplasia), making the enamel thinner and more prone to decay.Comparisons between Gracile and Robust Australopiths
EARLY TOOL USE AND TECHNOLOGY
Early Stone Age Technology (ESA)
The Early Stone Age (ESA) marks the beginning of recognizable technology as made by our human ancestors. Stone-tool (or lithic) technology is defined by the fracturing of rocks and the manufacture of tools through a process called knapping. The Stone Age lasted for more than 3 million years and is broken up into chronological periods called the Early (ESA), Middle (MSA), and Later Stone Ages (LSA). Each period is further broken up in different techno-complexes, as explained below. This section focuses on the earliest tools associated with ESA. The ESA spanned the largest technological time period of human innovation from over 3 million years ago to around 300,000 years ago and is associated almost entirely with hominin species prior to modern Homo sapiens. As the ESA advanced, stone tool makers (known as knappers) began to change the ways they detached flakes and eventually were able to shape artifacts into functional tools. These advances in technology go together with the developments in human evolution and cognition, dispersal of populations across the African continent and the world, and climatic changes.
The earliest described archaeological period in which we start seeing stone tool technology.
Relating to stone (here to stone tools).
The fracturing of rocks for the manufacture of tools.
The piece knocked off of a stone core during the manufacture of a tool, which may be used as a stone tool.
In order to understand the ESA, it is important to consider some definitions. A techno-complex is a term encompassing multiple assemblages (collections of artifacts) that share similar traits in terms of artifact production and morphology. Not all assemblages are exactly the same within each techno-complex: one can have multiple phases and traditions at different sites (Lombard et al. 2012). However, there is an overarching commonality between them. Within stone tool assemblages, both flakes or cores (the rocks from which flakes are removed) are used as tools. Large Cutting Tools (LCTs) are tools that are shaped to have functional edges. It is important to note that the information presented here is a small fraction of what is known about the ESA, and there are many ongoing debates and discoveries within the archaeological discipline.
A term encompassing multiple assemblages that share similar traits in terms of artifact production and morphology.
A collection of artifacts demonstrating a pattern. Often pertaining to a site or region.
The remains of a rock that has been flaked or knapped.
A tool that is shaped to have functional edges.
Currently, the oldest known stone tools, which form the techno-complex the Lomekwian, date to 3.3 mya (Harmand et al. 2015; Toth 1985). They were found at a site called Lomekwi 3 in Kenya. This techno-complex is the most recently defined and pushed back the oldest known date for lithic technology. There is only one known site thus far and, due to the age of the site, it is associated with species prior to Homo, such as Kenyanthropus platyops. Flakes were produced through indirect percussion, whereby the knappers held a rock and hit it against another rock resting on the ground. The pieces are very chunky and do not display the same fracture patterns as seen in later techno-complexes. Lomekwian knappers likely aimed to get a sharp-edged piece on a flake, which would have been functional, although the specific function is currently unknown.
Stone tool use, however, is not only understood through the direct discovery of the tools. Cut marks on fossilized animal bones may illuminate the functionality of stone tools. In one controversial study in 2010, researchers argued that cut marks on a pair of animal bones from Dikika (Ethiopia), dated to 3.4 mya, were from stone tools. The discoverers suggested that they be more securely associated, temporally, with Au. afarensis. However, others have noted that these marks are consistent with teeth marks from crocodiles and other carnivores.
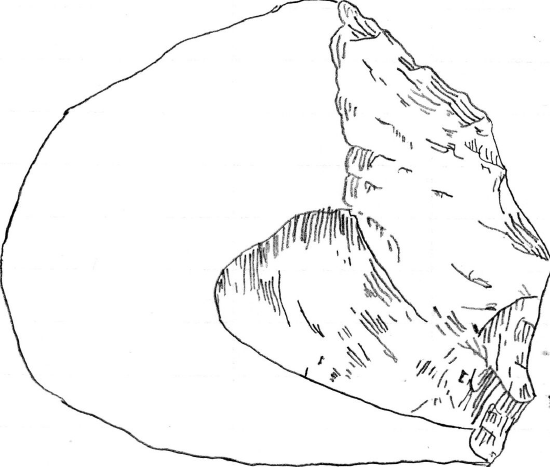
The Oldowan techno-complex is far more established in the scientific literature (Leakey 1971). It is called the Oldowan because it was originally discovered in Olduvai Gorge, Tanzania, but the oldest assemblage is from Gona in Ethiopia, dated to 2.6 mya (Semaw 2000). The techno-complex is defined as a core and flake industry. Like the Lomekwian, there was an aim to get sharp-edged flakes, but this was achieved through a different production method. Knappers were able to actively hold or manipulate the core being knapped, which they could directly hit using a hammerstone. This technique is known as free-hand percussion, and it demonstrates an understanding of fracture mechanics. It has long been argued that the Oldowan hominins were skillful in tool manufacture.
Lower Paleolithic, the earliest stone tool culture consisting of simple flakes and choppers.
Because Oldowan knapping requires skill, earlier researchers have attributed these tools to members of our genus, Homo. However, some have argued that these tools are in more direct association with hominins in the genera described in this chapter (Figure \(\PageIndex{14}\)).
Invisible Tool Manufacture and Use
The vast majority of our understanding of these early hominins comes from fossils and reconstructed paleoenvironments. It is only from 3 mya when we can start “looking into their minds” and lifestyles by analyzing their manufacture and use of stone tools. However, the vast majority of tool use in primates (and, one can argue, in humans) is not with durable materials like stone. All of our extant great ape relatives have been observed using sticks, or leaves, or other materials for some secondary purpose (to wade across rivers, to “fish” for termites, or to absorb water for drinking). It is possible that the majority of early hominin tool use and manufacture may be invisible to us because of this preservation bias.
REFERENCES
Alemseged, Z., F. Spoor, W. H. Kimbel, R. Bobe, D. Geraads, D. Reed, and J. G. Wynn. 2006. “A Juvenile Early Hominin Skeleton from Dikika, Ethiopia.” Nature 443 (7109): 296–301.
Asfaw, B., T. White, O. Lovejoy, B. Latimer, S. Simpson, and G. Suwa. 1999. “Australopithecus garhi: A New Species of Early Hominid from Ethiopia.” Science 284 (5414): 629–635.
Berger, L. R., D. J. De Ruiter, S. E. Churchill, P. Schmid, K. J. Carlson, P. H. Dirks, and J. M. Kibii. 2010. “Australopithecus sediba: A New Species of Homo-like Australopith from South Africa.” Science 328 (5975): 195–204.
Brain, C. K. 1967. “The Transvaal Museum’s Fossil Project at Swartkrans.” South African Journal of Science 63 (9): 378–384.
Broom, R. 1938a. “More Discoveries of Australopithecus.” Nature 141 (1): 828–829.
Broom, R. 1938b. “The Pleistocene Anthropoid Apes of South Africa.” Nature 142 (3591): 377–379.
Broom, R.. 1947. “Discovery of a New Skull of the South African Ape-Man, Plesianthropus.” Nature 159 (4046): 672.
Broom, R.. 1950. “The Genera and Species of the South African Fossil Ape-Man.” American Journal of Physical Anthropology 8 (1): 1–14.
Brunet, M., A. Beauvilain, Y. Coppens, E. Heintz, A. H. Moutaye, and D. Pilbeam. 1995. “The First Australopithecine 2,500 Kilometers West of the Rift Valley (Chad).” Nature 378 (6554): 275–273.
Clarke, R. J. 1998. “First Ever Discovery of a Well-Preserved Skull and Associated Skeleton of Australopithecus.” South African Journal of Science 94 (10): 460–463.
Clarke, R.J. 2013. “Australopithecus from Sterkfontein Caves, South Africa.” In The Paleobiology of Australopithecus, edited by K.E. Reed, J.G. Fleagle, and R.E. Leakey, 105–123. Netherlands: Springer.
Clarke, R. J. and K. Kuman. 2019. “The Skull of StW 573, a 3.67 Ma Australopithecus Prometheus Skeleton from Sterkfontein Caves, South Africa.” Journal of Human Evolution 134: 102634.
Clarke, R. J. and P. V. Tobias. 1995. Sterkfontein Member 2 Foot Bones of the Oldest South African Hominid. Science 269 (5223): 521–524.
Constantino, P. J. and B. A. Wood. 2004. Paranthropus Paleobiology. In Miscelanea en Homenae a Emiliano Aguirre, volumen III: Paleoantropologia, edited by E.G. Pérez and S.R. Jara, 136–151. Alcalá de Henares: Museo Arqueologico Regional.
Constantino, P. J. and B. A. Wood. 2007. “The Evolution of Zinjanthropus boisei.” Evolutionary Anthropology: Issues, News, and Reviews 16 (2): 49–62.
Dart, R. A. 1925. “Australopithecus africanus, the Man-Ape of South Africa.” Nature 115: 195–199.
Dirks, P. H., J. M. Kibii, B. F. Kuhn, C. Steininger, S. E. Churchill, J. D. Kramers, and G. C. King. 2010. “Geological Setting and Age of Australopithecus sediba from Southern Africa.” Science 328 (5975): 205–208.
Grine, F. E. 1988. “New Craniodental Fossils of Paranthropus from the Swartkrans Formation and Their Significance in “Robust” Australopithecine Evolution.” In Evolutionary History of the “Robust” Australopithecines, edited by F. E. Grine, 223–243. New York: Aldine de Gruyter.
Haile-Selassie, Y., L. Gibert, S. M. Melillo, T. M. Ryan, M. Alene, A. Deino, G. Scott, and B. Z. Saylor. 2015. “New Species from Ethiopia Further Expands Middle Pliocene Hominin Diversity.” Nature 521 (7553): 432–433.
Haile-Selassie, Y., S. M. Melillo, A. Vazzana, S. Benazzi, and T. M. Ryan. 2019. “A 3.8-Million-Year-Old Hominin Cranium from Woranso-Mille, Ethiopia.” Nature 573 (7773): 214-219.
Harmand, S., J. E. Lewis, C. S. Feibel, C. J. Lepre, S. Prat, A. Lenoble, X. Boë, et al. 2015. “3.3-million-year-old Stone Tools from Lomekwi3, West Turkana, Kenya.” Nature 521(7552): 310–316.
Hay, R. L. 1990. “Olduvai Gorge: A Case History in the Interpretation of Hominid Paleoenvironments.” In East Africa: Establishment of a Geologic Framework for Paleoanthropology, edited by L. Laporte, 23–37. Boulder: Geological Society of America.
Hay, R. L. and M. D. Leakey. 1982. “The Fossil Footprints of Laetoli.” Scientific American 246 (2): 50–57.
Hlazo, N. 2015. “Paranthropus: Variation in Cranial Morphology.” Honours thesis, Archaeology Department, University of Cape Town, Cape Town.
Hlazo, N. 2018. “Variation and the Evolutionary Drivers of Diversity in the Genus Paranthropus.” Master’s thesis, Archaeology Department, University of Cape Town, Cape Town.
Johanson, D. C., T. D. White, and Y. Coppens. 1978. “A New Species of the Genus Australopithecus (Primates: Hominidae) from the Pliocene of East Africa.” Kirtlandia 28: 1–14.
Kimbel, W. H. 2015. “The Species and Diversity of Australopiths. In Handbook of Paleoanthropology, Second Edition, edited by T. Hardt, 2071–2105. Berlin: Springer.
Kimbel, W. H. and L. K. Delezene. 2009. “‘Lucy’ Redux: A Review of Research on Australopithecus afarensis.” American Journal of Physical Anthropology 140 (S49): 2–48.
Leakey, L. S. B. 1959. “A New Fossil Skull from Olduvai.” Nature 184 (4685): 491–493.
Leakey, M. 1971. Olduvai Gorge, Vol. 3. Cambridge: Cambridge University Press.
Leakey, M. G., C. S. Feibel, I. McDougall, and A. Walker. 1995. “New Four–million-year-old Hominid Species from Kanapoi and Allia Bay, Kenya.” Nature 376 (6541): 565–571.
Leakey, M. D. and R. L. Hay. 1979. “Pliocene Footprints in the Laetoli Beds at Laetoli, Northern Tanzania.” Nature 278 (5702): 317–323.
Leakey, M. G., F. Spoor, F. H. Brown, P. N. Gathogo, C. Kiarie, L. N. Leakey, and I. McDougall. 2001. “New Hominin Genus from Eastern Africa Shows Diverse Middle Pliocene Lineages.” Nature 410 (6827): 433–440.
Lebatard, A. E., D. L. Bourlès, P. Duringer, M. Jolivet, R. Braucher, J. Carcaillet, and A. Likius. 2008. “Cosmogenic Nuclide Dating of Sahelanthropus tchadensis and Australopithecus bahrelghazali: Mio-Pliocene Hominids from Chad.” Proceedings of the National Academy of Sciences 105 (9): 3226–3231.
Lee-Thorp, J. 2011. “The Demise of ‘Nutcracker Man.’” Proceedings of the National Academy of Sciences 108 (23): 9319–9320.
Lombard, M., L. Wadley, J. Deacon, S. Wurs, I. Parsons, M. Mohapi, J. Swart, and P. Mitchell. 2012. “South African and Lesotho Stone Age Sequence Updated.” The South African Archaeological Bulletin 67 (195): 123–144.
McHenry, H. M. 2009. “Human Evolution.” In Evolution: The First Four Billion Years, edited by M. Ruse and J. Travis, 256–280. Cambridge: The Belknap Press of Harvard University Press.
Patterson, B. and W. W. Howells. 1967. “Hominid Humeral Fragment from Early Pleistocene of Northwestern Kenya.” Science 156 (3771): 64–66.
Pickering, R. and J. D. Kramers. 2010. “Re-appraisal of the Stratigraphy and Determination of New U-Pb Dates for the Sterkfontein Hominin Site.” Journal of Human Evolution 59 (1): 70–86.
Rak, Y. 1983. The Australopithecine Face. New York: Academic Press.
Rak, Y. 1988. “On Variation in the Masticatory System of Australopithecus boisei.” In Evolutionary History of the “Robust” Australopithecines, edited by M. Ruse and J. Travis, 193–198. New York: Aldine de Gruyter.
Semaw, S. 2000. “The World’s Oldest Stone Artefacts from Gona, Ethiopia: Their Implications for Understanding Stone Technology and Patterns of Human Evolution Between 2.6 Million Years Ago and 1.5 Million Years Ago.” Journal of Archaeological Science 27: 1197–1214.
Spoor, F. 2015. “Palaeoanthropology: The Middle Pliocene Gets Crowded.” Nature 521 (7553): 432– 433.
Strait, D. S., F. E. Grine, and M. A. Moniz. 1997. A Reappraisal of Early Hominid Phylogeny.” Journal of Human Evolution 32 (1): 17–82.
Thackeray, J. F., Braga, J. Treil, N. Niksch, and J. H. Labuschagne. 2002. “‘Mrs. Ples’ (Sts 5) from Sterkfontein: An Adolescent Male?” South African Journal of Science 98 (1–2): 21–22.
Thackeray, J. F. 2000. “‘Mrs. Ples’ from Sterkfontein: Small Male or Large Female?” The South African Archaeological Bulletin, 55: 155–158.
Toth, N. 1985. “The Oldowan Reassessed.” Journal of Archaeological Science 12(2): 101–120.
Walker, A. C., R. E. Leakey, J. M. Harris, and F. H. Brown. 1986. “2.5-my Australopithecus boisei from West of Lake Turkana, Kenya.” Nature 322 (6079): 517–522.
Walker, A. C. and R. E. Leakey. 1988. “The Evolution of Australopithecus boisei.” In Evolutionary History of the “Robust” Australopithecines, edited by F. E. Grine, 247–258. New York: Aldine de Gruyter.
Ward, C., M. Leakey, and A. Walker. 1999. “The New Hominid Species Australopithecus anamensis.” Evolutionary Anthropology 7(6): 197–205.
White, T. D. 1988. “The Comparative Biology of ‘Robust’ Australopithecus: Clues from Content.” In Evolutionary History of the “Robust” Australopithecines, edited by F. E. Grine, 449–483. New York: Aldine de Gruyter.
Wood, B. A. 2010. “Reconstructing Human Evolution: Achievements, Challenges, and Opportunities.” Proceedings of the National Academy of Sciences 10(2): 8902–8909.
Wood, B. A. and E. K. Boyle. 2016. “Hominin Taxic Diversity: Fact or Fantasy?” Yearbook of Physical Anthropology 159 (S61): 37–78.
Wood, B. and K. Schroer. 2017. “Paranthropus: Where Do Things Stand?” In Human Paleontology and Prehistory, edited by A. Marom and E. Hovers, 95–107. New York: Springer, Cham.
FIGURE ATTRIBUTIONS
Figure \(\PageIndex{1}\) Paranthropus bonsei compared to Homo sapiens by Constantino, Paul J. is used under a CC BY-SA 4.0 License.
Figure \(\PageIndex{2}\) Australopithecus anamensis: KNM-KP 29281 occlusal view by eFossils is copyrighted and used for noncommercial purposes as outlined by eFossils.
Figure \(\PageIndex{3}\) Derived from Australopithecus afarensis, “Lucy,” adult female. Reconstruction based on AL-288-1 by artist John Gurche, front view close-up and Australopithecus afarensis, adult male. Reconstruction based on AL444-2 by John Gurche by Smithsonian [exhibit: Reconstructed Faces, What does it mean to be human?] is copyrighted and used for educational and noncommercial purposes as outlined by the Smithsonian.
Figure \(\PageIndex{4}\) Lucy blackbg (AL 288-1, Australopithecus afarensis, cast from Museum national d’histoire naturelle, Paris) by 120 is used under a CC BY-SA 3.0 License.
Figure \(\PageIndex{5}\) Derived from Kenyanthropus platyops KNM WT 40000 anterior view, Kenyanthropus platyops KNM WT 40000 superior view, Kenyanthropus platyops KNM WT 40000 lateral left view, Kenyanthropus platyops KNM WT 40000 inferior view by eFossils is copyrighted and used for noncommercial purposes as outlined by eFossils.
Figure \(\PageIndex{6}\) Derived from australopithecus africanus: Taung 1 anterior view and australopithecus africanus: Taung 1 lateral right view by eFossils is copyrighted and used for noncommercial purposes as outlined by eFossils.
Figure \(\PageIndex{7}\) Australopithecus africanus. Reconstruction based on STS 5 by John Gurche by Smithsonian [exhibit: Reconstructed Faces, What does it mean to be human?] is copyrighted and used for educational and noncommercial purposes as outlined by the Smithsonian.
Figure \(\PageIndex{8}\) Derived from Australopithecus africanus Sts 5 anterior view , Australopithecus africanus Sts 5 posterior view, Australopithecus africanus Sts 5 superior view, Australopithecus africanus Sts 5 lateral right view by eFossils is copyrighted and used for noncommercial purposes as outlined by eFossils.
Figure \(\PageIndex{9}\) Australopithecus sediba, photo by Brett Eloff, courtesy Profberger and Wits University is used under a CC BY-SA 4.0 License.
Figure \(\PageIndex{10}\) Derived from Paranthropus aethiopicus: KNM-WT 17000 anterior view; Paranthropus aethiopicus: KNM-WT 17000 lateral right view; Paranthropus aethiopicus: KNM-WT 17000 superior view; Paranthropus aethiopicus: KNM-WT 17000 posterior view; Paranthropus aethiopicus: KNM-WT 17000 inferior view by eFossils is copyrighted and used for noncommercial purposes as outlined by eFossils.
Figure \(\PageIndex{11}\) Paranthropus boisei, male. Reconstruction based on OH 5 and KNM-ER 406 by John Gurche by Smithsonian [exhibit: Reconstructed Faces, What does it mean to be human?] is copyrighted and used for educational and noncommercial purposes as outlined by the Smithsonian.
Figure \(\PageIndex{12}\) Derived from Paranthropus boisei: OH 5 anterior view; Paranthropus boisei: OH 5 inferior view; and Paranthropus boisei: OH 5 posterior view; by eFossils is copyrighted and used for noncommercial purposes as outlined by eFossils.
Figure \(\PageIndex{13}\) Derived from Paranthropus robustus: SK 48 anterior view; Paranthropus robustus: SK 48 superior view; Paranthropus robustus: SK 48 inferior view; Paranthropus robustus: SK 48 lateral left view by eFossils is copyrighted and used for noncommercial purposes as outlined by eFossils.
Figure \(\PageIndex{14}\) Olduwan Industry Chopper 2 by Emmyanne29 is used under a CC0 1.0 License.