
10.2: Homo Habilis
- Page ID
- 158781
THE EARLIEST MEMBERS OF OUR GENUS
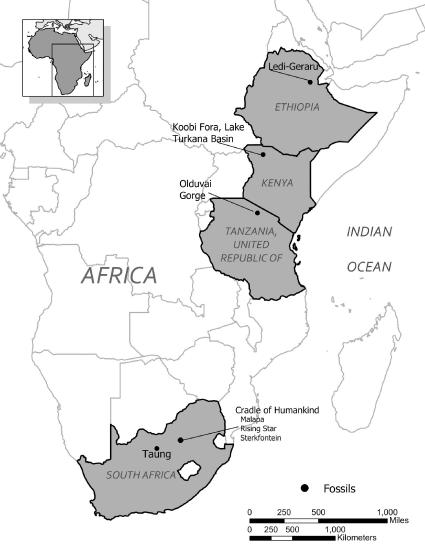 Figure \(\PageIndex{1}\): Map showing major sites where Homo habilis fossils have been found.
Figure \(\PageIndex{1}\): Map showing major sites where Homo habilis fossils have been found.Discovery and Naming
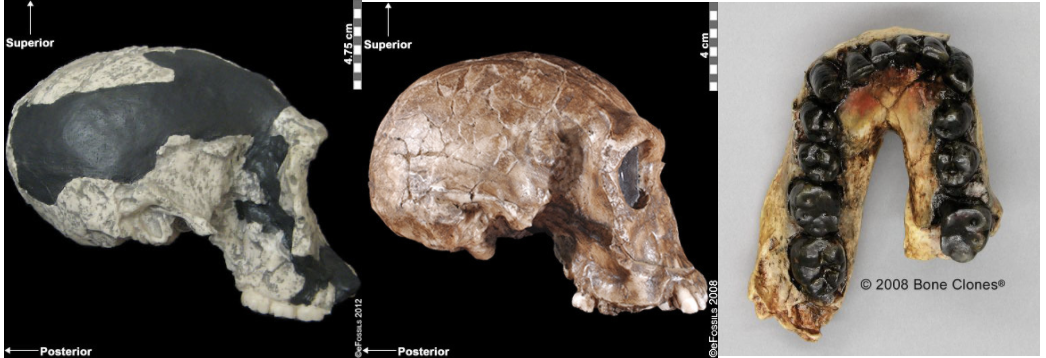 Figure \(\PageIndex{2}\): Homo habilis fossil specimens. From left to right they are: OH-24 (found at Olduvai Gorge), KNM-ER-1813 (from Koobi Fora, Kenya), and the jaw of OH-7, which was the type specimen found in 1960 at Olduvai Gorge, Tanzania.
Figure \(\PageIndex{2}\): Homo habilis fossil specimens. From left to right they are: OH-24 (found at Olduvai Gorge), KNM-ER-1813 (from Koobi Fora, Kenya), and the jaw of OH-7, which was the type specimen found in 1960 at Olduvai Gorge, Tanzania.Controversies over Classification of Homo habilis
How Many Species of Homo habilis?
The capability of an organism to modify its phenotype during development in response to environmental cues.
Homo habilis: Homo or Australopithecus?
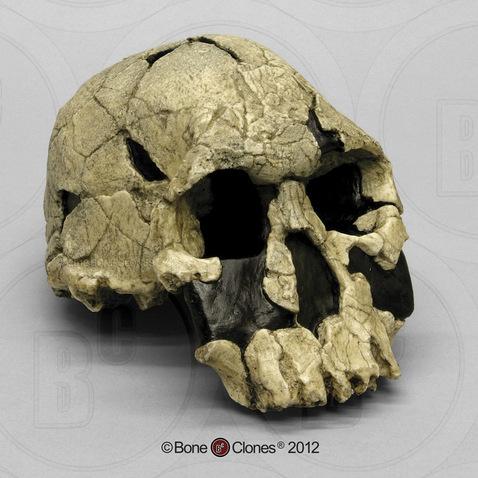 Figure \(\PageIndex{3}\): Cast of the Homo habilis cranium KNM-ER-1470. This cranium has a wide, flat face, larger brain size, and larger teeth than other Homo habilis fossils, leading some scientists to give it a separate species name, Homo rudolfensis.
Figure \(\PageIndex{3}\): Cast of the Homo habilis cranium KNM-ER-1470. This cranium has a wide, flat face, larger brain size, and larger teeth than other Homo habilis fossils, leading some scientists to give it a separate species name, Homo rudolfensis.HOMO HABILIS CULTURE AND LIFEWAYS
Early Stone Tools
The larger brains and smaller teeth of early Homo are linked to a different adaptive strategy than that of earlier hominins—one dependent on modifying rocks to make stone tools and exploit new food sources. Based on what we know from nonhuman-primate tool use, it is assumed that all hominins used tools of some sort. For example, australopithecines could have used digging sticks to extract the roots and tubers that were part of some species’ diets (though tools made from perishable material would leave no trace). As discussed in the previous chapter, stone tools almost certainly predated Homo habilis (possibly by Australopithecus garhi or the species responsible for the tools from Kenya dating to 3.7 million years ago). However, stone tools become more frequent at sites dating to about 2 million years ago, the time of Homo habilis (Roche, Blumenschine, and Shea 2009). This suggests that these hominins were increasingly reliant on stone tools to make a living.
Stone tools are assigned a good deal of importance in the study of human origins. Studying the form of the tools, the raw materials selected, and how they were made and used can provide insight into the thought processes of early humans and how they modified their environment in order to survive. Paleoanthropologists have traditionally classified collections of stone tools into industries, based on their form and mode of manufacture. There is not an exact correspondence between a tool industry and a hominin species; however, some general associations can be made between tool industries and particular hominins, locations, and time periods. The names for the four primary tool industries in human evolution (from oldest to most recent) are the Oldowan, Acheulean, Mousterian, and Upper Paleolithic.
The oldest stone tool industry is the Oldowan, named after the site of Olduvai Gorge where the tools were first discovered. The time period of the Oldowan is generally considered to last from about 2.5 mya to 1.6 mya. The tools of this industry are described as “flake and chopper” tools—the choppers consisting of stone cobbles with a few flakes struck off them (Figure \(\PageIndex{4}\)). To a casual observer, these tools might not look much different from randomly broken rocks. However, they are harder to make than their crude appearance suggests. The rock selected as the core must be struck by the rock serving as a hammerstone at just the right angle so that one or more flat flakes are removed. This requires selecting rocks that will fracture predictably instead of chunking, as well as the ability to plan ahead and envision the steps needed to create the finished product. The process leaves both the core and the flakes with sharp cutting edges that can be used for a variety of purposes.
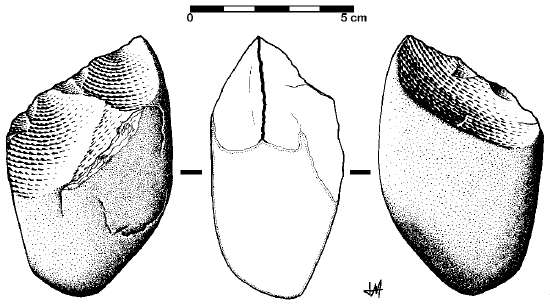 Figure \(\PageIndex{4}\): Drawing of an Oldowan-style tool. This drawing shows a chopper; the flakes removed from the cores functioned as cutting tools.
Figure \(\PageIndex{4}\): Drawing of an Oldowan-style tool. This drawing shows a chopper; the flakes removed from the cores functioned as cutting tools.Stone Tool Use and the Diet of Early Homo
What were the hominins doing with the tools? One key activity seems to have been butchering animals. Animal bones with cutmarks start appearing at sites with Oldowan tools. Studies of animal bones at the site show leg bones are often cracked open, suggesting that they were extracting the marrow from the bone cavities. It is interesting to consider whether the hominins hunted these animals or acquired them through other means. The butchered bones come from a variety of African mammals, ranging from small antelope to animals as big as wildebeest and elephants! It is difficult to envision slow, small-bodied Homo habilis with their Oldowan tools bringing down such large animals. One possibility is that the hominins were scavenging carcasses from lions and other large cats. Paleoanthropologist Robert Blumenschine has evaluated the scavenging hypothesis by directly observing the behavior of present-day animal carnivores and scavengers on the African savanna. From this, he inferred that there were scavenging opportunities for Plio-pleistocene hominins. When lions abandon a kill after eating their fill, scavenging animals arrive almost immediately to pick apart the carcass. By the time the slow-footed hominins arrived on the scene, the carcass would be mostly stripped of meat. However, if hominins could use stone tools to break into the leg bone cavities, they could get to the marrow, a fatty, calorie-dense source of protein (Blumenschine et al 1987).
Reconstructing activities that happened millions of years ago is obviously a difficult undertaking, and there is an active debate among anthropologists about whether scavenging or hunting was more commonly practiced during this time. Regardless of how they were acquiring the meat, all these activities suggest an important dietary shift from the way that the australopithecines were eating. The Oldowan toolmakers were exploiting a new ecological niche that provided them with more protein and calories. And it was not just limited to meat-eating—stone tool use could have made available numerous other subsistence opportunities. A study of microscopic wear patterns on a sample of Oldowan tools indicates that they were used for processing plant materials such as wood, roots or tubers, and grass seeds and stems (Lemorini et al. 2014). In fact, it has been pointed out that the Oldowan toolmakers’ cutting ability (whether for the purposes of consuming meat and plants or for making tools, shelters or clothing) represents a new and unique innovation, never seen before in the natural world! (Roche, Blumenschine, and Shea 2009).
Overall, increasing use of stone tools allowed hominins to expand their ecological niche and exert more control over their environment. As we’ll see shortly, this pattern continued and became more pronounced with Homo erectus.
REFERENCES
Anton, Susan C., Richard Potts, and Leslie C. Aiello. 2014. “Evolution of Early Homo: An Integrated Biological Perspective.” Science 345 (6192) doi: 10.1126/science.1236828.
Blumenschine, Robert, Henry T. Bunn, Valerius Geist, Fumiko Ikawa-Smith, Curtis W. Marean, Anthony G. Payne, John Tooby, J. Nikolaas, and Van Der Merwe. 1987. “Characteristics of an Early Hominid Scavenging Niche [and Comments and Reply].” Current Anthropology 28 (4): 383–407.
Leakey, Louis S. B., Phillip V. Tobias, and John R. Napier. 1964. “A New Species of Genus Homo from Olduvai Gorge.” Nature 202: 308–312.
Lemorini, Cristina, Thomas W. Plummer, David R. Braun, Alyssa N. Crittenden, Peter W. Ditchfield, Laura C. Bishop, Fritz Hertel, James S. Oliver, Frank W. Marlowe, Margaret J. Schoeninger, and Richard Potts. 2014. “Old Stones’ Song: Use-wear Experiments and Analysis of the Oldowan Quartz and Quartzite Assemblage from Kanjera South (Kenya).” Journal of Human Evolution 72: 10–25.
Roche, Helene, Robert J. Blumenschine, and John J. Shea. 2009. “Origins and Adaptations of Early Homo: What Archeology Tells Us.” In The First Humans: Origin and Early Evolution of the Genus Homo, edited by Frederick E. Grine, John G. Fleagle, and Richard E. Leakey, 135-147. New York: Springer.
Ruff, Christopher. 2009. “Relative Limb Strength and Locomotion in Homo habilis.” American Journal of Physical Anthropology 138 (1): 90–100.
Villmoare, Brian, William H. Kimbel, Chalachew Seyoum, Christopher J. Campisano, Erin N. DiMaggio, John Rowan, David R. Braun, J. Ramón Arrowsmith, and Kaye E. Reed. 2015. “Early Homo at 2.8 Ma From Ledi-Geraru, Afar, Ethiopia.” Science 347 (6228): 1352–1355.
Wood, Bernard. 2014. “Human Evolution: Fifty Years after Homo habilis.” Nature 508 (7494): 31–33.
Wood, Bernard, and Mark Collard. 1999. “The Changing Face of Genus Homo.” Evolutionary Anthropology 8 (6): 195–207.
FIGURE ATTRIBUTIONS
Figure \(\PageIndex{1}\) Homo habilis site map original to Explorations: An Open Invitation to Biological Anthropology by Chelsea Barron at GeoPlace, California State University, Chico is under a CC BY-NC 4.0 License.
Figure \(\PageIndex{2}\) Derivitive of Homo habilis: OH 24 lateral right view; Homo habilis: KNM-ER 1813 lateral right view by eFossils is copyrighted and used for non-commercial purposes as outlined by eFossils; and, Homo habilis OH 7 Jaw by ©BoneClones is used by permission and available here under a CC BY-NC 4.0 License.
Figure \(\PageIndex{3}\) Homo rudolfensis Cranium KNM-ER 1470 by ©BoneClones is used by permission and available here under a CC BY-NC 4.0 License.
Figure \(\PageIndex{4}\) Chopping tool by José-Manuel Benito Álvarez is used under a CC BY-SA 2.5 License.
TABLE ATTRIBUTIONS
Table \(\PageIndex{1}\) Homo habilis table original to Explorations: An Open Invitation to Biological Anthropology is under a CC BY-NC 4.0 License.
Table \(\PageIndex{2}\) Summary features of Homo habilis original to Explorations: An Open Invitation to Biological Anthropology is under a CC BY-NC 4.0 License.