
5.3: Between-Neuron Communicaton- Synaptic Transmission
- Page ID
- 173128
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Learning Objectives
- Differentiate the components of a typical electrical and chemical synapse
- Describe ion channels, and what changes they undergo when neuron potentials are produced; what causes ion channels to change during synaptic transmission?
- Define ionotropic and metabotropic receptors and discuss in what ways they differ from one another in their effects during synaptic transmission
- Explain the steps in synaptic transmission from pre-synaptic neuron to post-synaptic neuron
- Describe how excitatory and inhibitory transmitters differ in their effects on post-synaptic neurons during synaptic transmission
- Discuss how layers of neurons, simulated by processing units in artificial neural networks, might produce psychological processes such as learning
Overview
Even before the action potential (i.e. "Nerve Impulse") has made it's way down to a neuron's axon terminals, the next form of neural communication has already begun. Thus, as Within-Neuron communication winds down, Between-Neuron (synaptic) Communication takes off in order to transmit the message to the next neuron (muscle or gland) in the circuit. In this section, we will explore the overlap of within- and between-neuron communication and examine the specifics of both electrical and chemical communication at the point of contact between neurons (i.e. synapses). The key structural elements of the synapse will be covered, including details on the receptors which receive the chemical messages known as neurotransmitters. All the steps of synaptic communication and their involvement in successful communication will be addressed.
Synapses
Recall that axons often split into several branches so that they can communicate with several other neurons. At the end of the axon's branches, the axon terminals typically form "synapses" with the dendritic spines or small protrusions on the dendrites of receiving neurons. Synapses form between the axon terminal of the presynaptic neuron (neuron sending the signal) and the postsynaptic neuron (neuron receiving the signal). See Figure \(\PageIndex{1}\)). Here we will focus specifically on synapses between the axon terminal of an axon and a dendritic spine; however, synapses can also form between the axon terminal of the presynaptic neuron and the postsynaptic neuron's soma, dendritic shaft, or the axon of another neuron.
A very small space called a synapse or synaptic gap/cleft exists between the pre-synaptic axon's terminal and the post-synaptic neuron's dendritic spine. To give you an idea of the size of the synaptic gap, a dime is 1.35 mm (millimeter) thick. There are 1,350,000 nm (nanometers) in the thickness of a dime. The synaptic gap is only about 5 nm wide. In the pre-synaptic terminal, there are storage packets made of fat called synaptic vesicles that package together the chemical messengers of neurons known as neurotransmitters (see Figure \(\PageIndex{1}\)). These vesicles protect neurotransmitters from breakdown and aid in their release from the presynaptic axon's terminal. Upon release into the synapse, the transmitters travel across the synapse to communicate a message to the postsynaptic neuron. They accomplish this by binding to a protein on the postsynaptic cell known as a receptor. More detail on the steps of synaptic transmission will be detailed below.
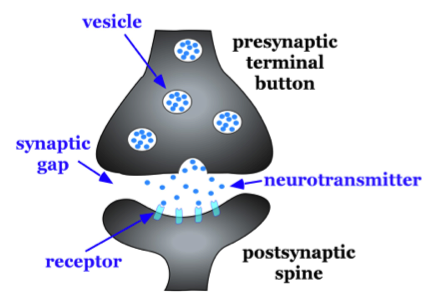
Figure \(\PageIndex{1}\): Basic characteristics of a typical synapse. Enlargement of the synapse between one of the axon terminal buttons (presynaptic terminal button) and one of the dendritic spines of a second neuron (postsynaptic spine).
SYNAPTIC TRANSMISSION
The between-neuron communication of chemical messages across the synaptic gap via neurotransmitters is one type of synaptic transmission. Synaptic transmission can also take place electrically without the need for the steps of chemical transmission. Synaptic transmission is central to the brain's capacity to process information, generate mental states, and produce adaptive behavior. Some synapses are purely electrical and make direct electrical connections between neurons, but, most synapses are chemical in nature and utilize neurotransmitters to transmit messages. The transmission of neural signals across chemical synapses is more complex than at electrical synapses and involves many steps.
Electrical Synapses
Electrical synapses are much less common than chemical synapses, but they are nevertheless distributed throughout the brain. In chemical synapses neurotransmitter is needed for communication between neurons, but for electrical synapses this is not the case. In electrical synapses current, ions, and molecules can flow between two neurons through direct physical connections that allow cytoplasm to flow between them. The physical connection between neurons with electrical synapses is in the form of large pore structures, called connexons, at gap junctions between such neurons. Communication between neurons with electrical synapses is faster than at chemical synapses which must go through more steps to transmit signals to another neuron. Therefore, electrical synapses are often found in neural systems that require rapid response such as defensive reflexes. Electrical synapses can communicate signals in both directions between neurons in contrast to chemical synapses which transmit messages in one direction, from pre-synaptic by transmitter release to post-synaptic neuron.
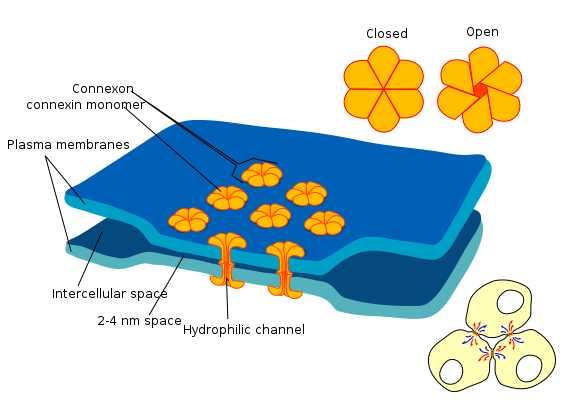
Figure \(\PageIndex{2}\): Gap junction at an electrical synapse. Two adjacent neurons with electrical synapse between them can communicate through hydrophilic channels Note that the gap between cell membranes of pre and post-synaptic neurons at electrical synapses is much smaller than the synaptic gap at chemical synapses which is about 10 times larger. (Image from Wikimedia Commons; File:Gap cell junction-en.svg; by Mariana Ruiz LadyofHats.https://commons.wikimedia.org/wiki/F...unction-en.svg; by Mariana Ruiz LadyofHats; public domain by its author, LadyofHats).
Chemical Synapses
Recall that in Within-Neuron Communication, several key charged particles (ions) are involved in maintaining the resting membrane potential and creating both post-synaptic and action potentials. At the end of this process, the action potential arrives at the axon terminals due to the exchange of sodium (Na+) and potassium (K+) ions that occurred along the length of the axon. Upon it's arrival at the terminal, the action potential triggers the opening of another ion channel which allows Calcium (Ca 2+) ions to enter. This entry of Ca 2+ ions triggers the release of chemical neurotransmitters into the synapse.
The release of neurotransmitter is actually the 3rd step of synaptic transmission. Prior to release, the transmitters have to first be synthesized and then stored. After neurotransmitters are released one of 3 other steps can occur. The transmitter can either bind to a receptor protein on the post-synaptic cell, be taken back into the presynaptic cell by a process known as reuptake, or metabolized (i.e. inactivation) by enzymes in the synapse. Reuptake and inactivation are both termination mechanisms for synaptic communication. In other words, they reduce or stop the between-neuron communication from continuing.
Steps of Synaptic Transmission
The process detailed in the previous section is known as the Steps of Synaptic Transmission. Depending on how you organize them, there are 5-6 of these steps as follows:
Synthesis, Storage, Release, Receptor Binding, Termination (Reuptake & Breakdown)
Let's explore these in a bit more detail.
Synthesis: There is a very large variety of neurotransmitters that act in the nervous system. Several of the major neurotransmitters will be described in detail in the Neurotransmitter section below. Of these, several are produced or synthesized by the enzymatic alterations of precursor amino acid molecules. In other words, certain amino acids from our diet are transformed into neurotransmitter molecules with the help of certain enzymes. Also, some neurotransmitters are manufactured in the cell body (soma) and transported to the axon terminal, while others (small ones) are synthesized in the axon terminal itself. Some of these synthesis pathways are more complicated than others, but all the resulting neurotransmitters share one thing in common. They are capable of communicating with other neurons and triggering or continuing a message.
Storage: With rare exception, neurotransmitters must be stored quickly after synthesis in order for them to be successfully released into the synapse. Structures made of the same fatty substance as the neuronal membrane are known as synaptic vesicles and serve to protect neurotransmitters from enzymatic breakdown and aid in their release to the synapse. As described previously, the the arrival of the action potential at the axon terminal causes the entry of Ca 2+ ions which trigger the release of neurotransmitters into the synapse. Well, the Ca 2+ ions actually interact with the synaptic vesicles and, essentially pushes them to the end of the terminal where they merge with the presynaptic membrane. This leads to the next step of neurotransmitter release.
Release: The release of neurotransmitter into the synapse is a critical step in the ultimate transmission of a message to the next neuron, muscle, or gland. As described above, it is the interaction of Ca 2+ ions with the synaptic vesicles that results in the release of the thousands of neurotransmitter molecules stored within them. Remember that the nerve cell releasing the neurotransmitter, the sender neuron, is known technically as the presynaptic neuron. The neuron receiving the neurotransmitter, the receiver cell, is called the postsynaptic neuron. The release transmitter must make it to the postsynaptic neuron in order to acheive the next step of message transmission. However, there are two other "termination" steps that often stop it from being successful.
Figure \(\PageIndex{9}\): A synapse. Synaptic vesicles release neurotransmitters (small yellow balls) which bind to the receptors (blue peg-like structures) on the postsynaptic membrane. Synaptic vesicles inside a presynaptic axon terminal button (axon ending) releasing neurotransmitter molecules onto receptors on a dendrite of a receiving (post-synaptic) neuron. (Image from Wikimedia Commons; File:Neurotransmitters.jpg; https://commons.wikimedia.org/wiki/F...ansmitters.jpg; by https://www.scientificanimations.com/; by https://www.scientificanimations.com/; licensed under the Creative Commons Attribution-Share Alike 4.0 International license).
Binding: If the released neurotransmitter avoids the termination mechanisms (see below), they may be able to bind to special sites on the membrane of the postsynaptic neuron. These special sites are proteins embedded in the membrane called receptors or postsynaptic receptor sites. They can be located anywhere on the neuron, but many are on the postsynaptic membrane of dendrites or dendritic spines of the receiving neuron. If their molecular shapes match the shapes of the neurotransmitter molecules, a molecular "lock and key" fit occurs which allows them to open (or close) chemically-gated ion channels. This change in the opening of ion channels is essentially the "message" and results in within-neuron electrical changes in voltage called postsynaptic potentials (PSPs). If there are enough excitatory post-synaptic potentials (EPSPs) to move the neuron past its activation threshold, an action potential will be produced and the whole communication process will continue.
There are more details on receptor binding in a special section toward the end of this section.
Termination Mechanisms - Inactivation
Special enzymes are present in the synapse and the neuron's themselves which are designed to metabolize and breakdown the neurotransmitter molecules. This enzymatic breakdown is known as inactivation and will stop the message in it's tracks. If neurotransmitter is not stored in vesicles and protected, it can be inactivated before it even gets released. If it is released, it can be inactivated by enzymes in the synapse before it has a chance to bind and interact with the post-synaptic neuron.
Termination Mechanisms - Reuptake
The other way the synaptic message can be reduced or terminated is via the process known as reuptake. Reuptake involves special proteins embedded in the PREsynaptic membrane which work very efficiently to clear the synapse of neurotransmitter by pulling it back inside the axon terminal. This is a type of recycling mechanism which reduces synaptic transmission and allows those transmitters to potentially be used again.
Summary of Steps of Synaptic Transmission
These steps of synaptic transmission from synthesis to the eventual termination of the neurotransmitter comprise synaptic transmission. Most, if not all of our thinking, feeling, and behaving rely on healthy function of this type of communication. Many psychological and physical issues relate to problems or dysregulation of this process. In addition, many psychoactive drugs (drugs that alter mind and/or behavior) such as nicotine, LSD, psilocybin, marijuana, etc. produce their effects by blocking or activating specific receptor sites. Other psychoactives produce their effects by altering one of the other steps of synaptic transmission, such as release or reuptake.
Here's a somewhat simplified but useful summary of the steps in synaptic transmission at synapses:
Figure \(\PageIndex{4}\): Image on left shows two neurons communicating with one another (synaptic transmission). The neuron at the top is the sender or pre-synaptic neuron. The neuron on the bottom is the receiver or post-synaptic neuron. In the image on the left of the figure, notice the small box at the synapse. This box is enlarged in the image on the right side of the figure to reveal details of the synapse and events there during synaptic transmission. The axon terminal of the pre-synaptic neuron is located at the top of the image on the right of the figure above. Transmitter molecules (small red squares) are in this axon terminal where they are stored in synaptic vesicles (yellow circles containing small red squares, i.e. transmitter molecules). (Image from Wikimedia Commons; File:Generic Neurotransmitter System.jpg; https://commons.wikimedia.org/wiki/F...ter_System.jpg; by NIDA(NIH); this work is in the public domain in the United States. Caption by Kenneth A. Koenigshofer, Ph.D.).
When a nerve impulse (action potential) reaches the axon terminal, calcium ions enter causing synaptic vesicles to move to the terminal membrane and release their transmitter molecules into the synapse. If they don't get inactivated by enzymes or undergo reuptake back into the terminal, the transmitter molecules may bind to post-synaptic receptors (green) causing specific ion channels ("doors") to open to specific ions. Ions (not shown in this figure) would then move through the ion channels causing a change in the voltage inside the post-synaptic neuron (either an EPSP or an IPSP). If trigger threshold (about negative 55 millivolts) is reached, an action potential is generated in the receiver neuron, which now becomes a sender neuron for the next cell in line (not shown in this figure). Inactivation of the used transmitter is the final step and is depicted in this figure by "enzyme degradation" (enzymatic destruction).
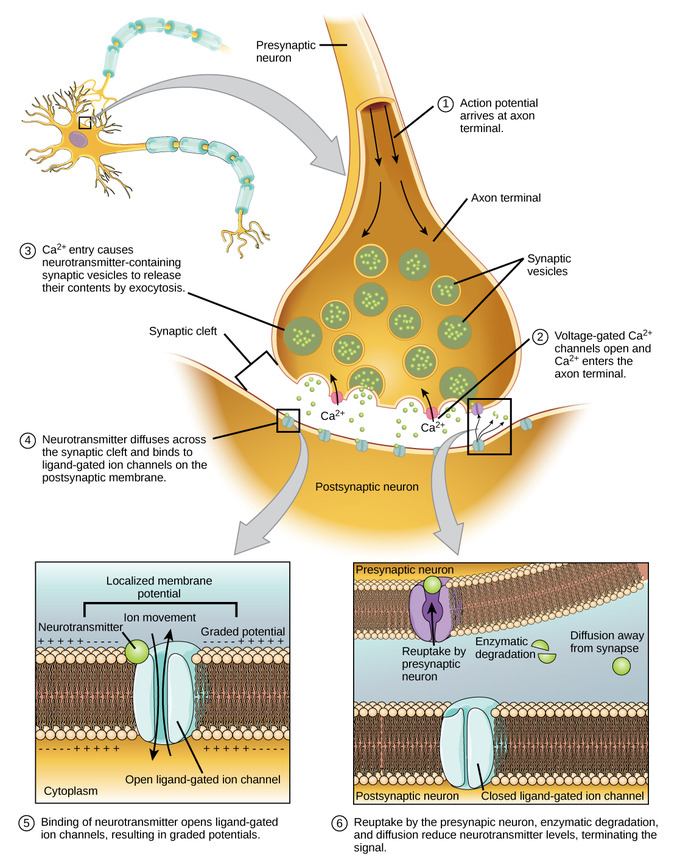
Figure \(\PageIndex{5}\): Communication at a chemical synapse: This illustrates most of the key steps of between-neuron, chemical communication that occur at the synapse. The initial step of "Synthesis" of neurotransmitter is not indicated, but would have occurred prior to storing the transmitter in the synaptic vesicles. Image adapted from: General Biology (Boundless), Chapter 35, The Nervous System; https://bio.libretexts.org/Bookshelv...gy_(Boundless); LibreTexts content is licensed by CC BY-NC-SA 3.0. Legal).
Communication at chemical synapses requires initial synthesis, storage, and then the release of neurotransmitters. When the depolarizing action potential arrives at the pre-synaptic axon terminal, voltage-gated Calcium (Ca2+) channels open and allow Ca2+ to enter the cell. The Ca2+ entry causes synaptic vesicles to fuse with the membrane and release neurotransmitter molecules into the synaptic cleft. The neurotransmitter can then do one of three things: It can either bind to a receptor, be taken back into the presynaptic terminal (reuptake), or be inactivated by enzymes in the synapse. If neurotransmitter diffuses across the synaptic cleft and binds to ligand-gated ion channels (channels opened by chemical transmitter) in the post-synaptic membrane, it will result in either a relatively small, localized depolarization or hyperpolarization of the post-synaptic neuron depending upon the type of receptor that it binds to. The local depolarizations are referred to as Excitatory Post Synaptic Potentials (EPSPs) and the local hyperpolarizations are Inhibitory Post Synaptic Potentials (IPSPs).
Receptor Binding
The binding of neurotransmitter to post-synaptic receptors is one of the most influential and dynamic steps of synaptic communication. All the other steps (synthesis, storage, release, reuptake, and breakdown) can be functioning, but without neurotransmitter binding, the chemical communication between neurons stops immediately. If the neurotransmitter does make it across the synapse and binds to a receptor, it can then produce one of a variety of effects depending on the type of receptor. Most neurotransmitters have a family of receptors which are able to bind them. Every system or circuitry of the brain contains particular transmitters and a subset of their receptor family.
To simplify, receptors vary based on two main characteristics: Type of post-synaptic effect that they cause and the speed of that effect.
The type of post-synaptic effect is determined by whether the receptor produces EPSPs or IPSPs, as described in the previous section. An "Excitatory" receptor will produce EPSPs, which will bring a neuron closer to the activation threshold for producing an action potential. An "Inhibitory" receptor will produce IPSPs which will make it more difficult for a neuron to reach the activation threshold. The particular type of receptor is determined by its molecular structure and the type of ions it will ultimately allow to flow into or out of the neuron. If the net effect is to cause the inside of the neuron to be less negative than it is at rest, it would be excitatory, and if it were to result in more negative charge inside the neuron it would be inhibitory.
In terms of the speed of effect, there are two major receptor subtypes: Ionotropic (Fast) and Metabotropic (Slow). In reality, both receptors produce very fast responses on the order of milliseconds, but metabotropics are slightly slower, as detailed below.
Ionotropic receptors, when activated by transmitter from a pre-synaptic neuron, cause ion channels to open, allowing ions, with their electrical charges, to move across the cell membrane of the receiving (post-synaptic) neuron, causing an EPSP or an IPSP (see above). These are fast acting receptors because the receptors are directly located on ion channels that open or close. In short, the receptor is an ion channel.
By contrast, metabotropic receptors (usually coupled with G-proteins; i.e. guanine nucleotide-binding proteins), when activated by transmitter from a pre-synaptic neuron, act indirectly and more slowly, using second messengers to produce a variety of metabolic effects to modulate cell activity. These effects include changes in gene transcription, regulation of proteins in the cell, release of Ca+ (calcium ions) within the cell, and effects on ion channels on the neuron's cell membrane (Sterling & Laughlin, 2015). Such modulation of neurons and synapses can be more long-lasting than effects of the activation of ionotropic receptors and may play an important role in cellular level mechanisms of learning and memory (Nadim and Bucher, 2014).
Figure \(\PageIndex{6}\): Comparison of Ionotropic and Metabotropic Post-Synaptic Receptors. The image at the top (a) shows ionotropic receptors which when activated by transmitter open ion channels immediately resulting in ion movement and an immediate response, a post-synaptic potential. The image at the bottom (b) shows metabotropic receptors which when activated by transmitter initiate a second messenger system. Second messengers can have a variety of effects including indirectly opening ion channels (Image from Wikimedia Commons; File:1226 Receptor Types.jpg; https://commons.wikimedia.org/wiki/F...ptor_Types.jpg; by OpenStax; licensed under the Creative Commons Attribution 4.0 International license).
After Receptor Binding
The binding of a neurotransmitter to its receptor is reversible, and for good reason. As long as it is bound to a receptor, a neurotransmitter continues to affect membrane potential, so it must eventually be removed from the synapse. The effects of the neurotransmitter generally lasts a few milliseconds before being terminated. If the used transmitter is not removed by the reuptake or inactivated by enzymes, it can cause over-activation of neurons, potentially leading to pathological mental states and behavior if enough synapses are affected. The two main types of neurotransmitter termination are: 1. Reuptake by astrocytes or the presynaptic axon terminal where the used transmitter can either be inactivated by enzymes or stored again, and 2. Inactivation by enzymes in the synaptic cleft. In some situations transmitter can also diffuse away from the synapse, thus stopping it's effects on the post-synaptic cell.
Types of Neurotransmitter
There are at least 60-100 neurotransmitters and probably many others yet to be discovered. The best known can be grouped into types based on their chemical structure.
Today, the majority of neuroscientists will tell you that most neurons release the same neurotransmitter from their axons—which is why you may see some neurons referred to as “dopaminergic” or “serotonergic,” releasing dopamine or serotonin, respectively. But new work in the field has uncovered that neurons are not fixed when it comes to the chemicals they release.
Some cells change the type of neurotransmitters they release depending on the circumstances, sometimes releasing up to five different kinds. Scientists call this phenomenon “neurotransmitter switching.”
Neurotransmitters, at the highest level, can be sorted into two types: small-molecule transmitters and neuropeptides. Small-molecule transmitters, like dopamine and glutamate, typically act directly on neighboring cells. The neuropeptides, small molecules like insulin and oxytocin, work more subtly, modulating, or adjusting, how cells communicate at the synapse. These powerful neurochemicals are at the center of neurotransmission, and, as such, are critical to human cognition and behavior.
Often, neurotransmitters are talked about as if they have a single role or function. Dopamine is a “pleasure chemical” and GABA is a “learning” neurotransmitter, even though glutamate also plays a major role in learning and memory formation. But neuroscientists are discovering they are multi-faceted and complex, working with and against each other to facilitate neural signaling across the cortex. Here is a list of some of the most common neurotransmitters discussed in neuroscience.
Amino acids neurotransmitters
- Glutamate (GLU). This is the most common and most abundant excitatory neurotransmitter. Glutamate has an important role in cognitive functions like thinking, learning and memory. Too much glutamate results in excitotoxicity, or the death of neurons due to stroke, traumatic brain injury, or amyotrophic lateral sclerosis (ALS), the debilitating neurodegenerative disorder better known Lou Gehrig’s disease. GLU is also important to learning and memory: long term potentiation (LTP), occurs in glutamatergic neurons in the hippocampus and cortex.
- Gamma-aminobutryic acid (GABA). GABA works to inhibit neural signaling. GABA is the most common inhibitory neurotransmitter in the nervous system, particularly in the brain. New research suggests that GABA helps lay down important brain circuits in early development. GABA also has a nickname: the “learning chemical.” Studies have found a link between the levels of GABA in the brain and whether or not learning is successful.
- Glycine. Glycine is the most common inhibitory neurotransmitter in the spinal cord and is involved in auditory processing, pain and metabolism.
Monoamines neurotransmitters
Monoamines neurotransmitters are involved in consciousness, cognition, attention and emotion.
- Serotonin (5HT). Serotonin is an inhibitory neurotransmitter. Serotonin helps regulate mood, sleep patterns, sexuality, anxiety, appetite and pain. Diseases associated with serotonin imbalance include seasonal affective disorder, anxiety, depression, fibromyalgia and chronic pain. Medications that regulate serotonin and treat these disorders include selective serotonin reuptake inhibitors (SSRIs) and serotonin-norepinephrine reuptake inhibitors (SNRIs), which increase the levels of transmitter by inhibiting their reuptake after they have done their work at the postsynaptic receptor sites. Serotonin (5HT), sometimes called the “calming chemical,” is best known for its mood modulating effects. A lack of 5HT has been linked to depression and related neuropsychiatric disorders. 5HT has also been implicated in facilitating memory, and, most recently, in decision-making behavior
- Histamine. Histamine regulates body functions including wakefulness, feeding behavior and motivation.
- Dopamine (DA). Dopamine is involved in the brain's reward system, feelings of pleasure, cognitive arousal, learning, focus of attention, concentration, memory, sleep, mood and motivation. Dopamine is often referred to as the “pleasure chemical” because it is released when mammals receive a reward in response to their behavior; that reward could be food, drugs, or sex. Diseases associated with dysfunctions of the dopamine system include Parkinson’s disease, schizophrenia, bipolar disease, and attention deficit hyperactivity disorder (ADHD). Many highly addictive drugs (cocaine, methamphetamines, amphetamines) act directly on the brain's dopamine circuits.
- Epinephrine. Epinephrine (also called adrenaline) and norepinephrine (see below) are responsible for the “fight-or-flight response” to fear and stress, activating the sympathetic nervous system (see Chapter 4).
- Norepinephrine (NE). Norepinephrine is both a hormone and a neurotransmitter. Some refer to it as noradrenalin. It has been linked to mood, arousal, vigilance, memory, and stress. Newer research has focused on its role in both post-traumatic stress disorder (PTSD) and Parkinson’s disease. Norepinephrine (noradrenaline) increases blood pressure, heart rate, alertness, arousal, attention and focus. Many medications (stimulants and depression medications) increase norepinephrine to improve focus or concentration to treat ADHD or to reduce symptoms of depression.
Peptide neurotransmitters
Peptides are short chains of amino acids.
- Endorphins. Endorphins are natural pain killers. They are natural opiate-like substances, similar in molecular properties to morphine. Release of endorphins reduces pain, and elevates mood. They are endogenous opiates released by the hypothalamus and pituitary gland during stress or pain.
Acetylcholine (ACh)
This excitatory neurotransmitter is found in both the central and peripheral nervous systems, specifically in the autonomic nervous system and in spinal motor neurons releasing acetylcholine onto skeletal muscles producing movements. It is also involved in memory, motivation, sexual desire, sleep and learning. Abnormalities in acetylcholine levels are associated with Alzheimer’s disease. But it also has other roles in the brain, including helping direct attention and playing a key role in facilitating neuroplasticity across the cortex.
Other Neurotransmitters
Neurochemicals like oxytocin and vasopressin are also classified as neurotransmitters. Made and released from the hypothalamus, they act directly on neurons and have been linked to pair-bond formation, monogamous behaviors, and drug addiction. Hormones like estrogen and testosterone can also work as neurotransmitters and influence synaptic activity.
Other neurotransmitter types include corticotropin-releasing factor (CRF), galanin, enkephalin, dynorphin, and neuropeptide Y. CRF, dynorphin, and neuropeptide Y have been implicated in the brain’s response to stress. Galanin, enkephalin, and neuropeptide Y are often referred to as “co-transmitters,” because they are released and then work in partnership with other neurotransmitters. Enkephalin, for example, is released with glutamate to signal the desire to eat and process rewards.
As neuroscientists are learning more about the complexity of neurotransmission, it’s clear that the brain needs these different molecules so it can have a greater range of flexibility and function.
Glia Release Neurotransmitters, Too
It was once believed that only neurons released neurotransmitters. New research, however, has demonstrated glia, the cells that make up the “glue” that fills the space between neurons to help support and maintain those cells, also have the power to release neurotransmitters into synapses. In 2004, researchers found that glial cells release glutamate into synapses in the hippocampus, helping synchronize signaling activity.
Astrocytes, a star-shaped glial cell, are known to release a variety of different neurotransmitters into the synapse to help foster synaptic plasticity, when required. Researchers are working diligently to understand the contributions of these different cell types–and the neurotransmitter molecules they release—on how humans think, feel, and behave.
Different Types of Receptors Activated by the Same Transmitter
There are different types of receptor for transmitters of a specific type. For example, for dopamine there are at least 5 types, D1 through D5 receptors, all for dopamine transmitter. Different receptor types for a specific transmitter may be localized in different parts of the brain and therefore may produce different effects and have different functions.
The function of each dopamine receptor type (Mishra, et al., 2018):
- D1: memory, attention, impulse control, regulation of renal (kidney) function, locomotion (movement)
- D2: locomotion, attention, sleep, memory, learning
- D3: cognition, impulse control, attention, sleep
- D4: cognition, impulse control, attention, sleep
- D5: decision making, cognition, attention, renin secretion (by the kidney)
Of particular interest, the mental disorder schizophrenia, characterized by disordered thought, hallucinations, and delusions, is associated with excess dopamine neuron activity in the brain. Some drug treatments for schizophrenia decrease activity primarily at D2 receptors.
Another example is acetylcholine transmitter. There are two distinct types of acetylcholine receptors affected by two different substances, either muscarine or nicotine. Those postsynaptic acetylcholine receptors that respond to muscarine are called muscarinic receptors. Those that respond to nicotine (in tobacco products, for example) are called nicotinic. Nicotinic receptors cause sympathetic postganglionic neurons and parasympathetic postganglionic neurons to fire and release their chemicals and cause skeletal muscle to contract. Muscarinic receptors are associated mainly with parasympathetic functions and stimulate receptors located in peripheral tissues (e.g., glands, smooth muscle). Acetylcholine transmitter activates all of these sites.
References
Bekkers, J. M. (2011). Pyramidal neurons. Current biology, 21(24), R975.
Brown, A. M., Arancillo, M., Lin, T., Catt, D. R., Zhou, J., Lackey, E. P., ... & Sillitoe, R. V. (2019). Molecular layer interneurons shape the spike activity of cerebellar Purkinje cells. Scientific reports, 9 (1), 1-19.
Churchland, P. M. (2013). Matter and consciousness. MIT press.
Fields, R. D., & Stevens-Graham, B. (2002). New insights into neuron-glia communication. Science, 298 (5593), 556-562.
Kirkcaldie, M. T. (2012). Neocortex. In The mouse nervous system (pp. 52-111). Academic Press.
Koenigshofer, K.A. (2011). Mind Design: The Adaptive Organization of Human Nature, Minds, and Behavior. Pearson Education, Boston.
Mishra A, Singh S, Shukla S. (2018). Physiological and Functional Basis of Dopamine Receptors and Their Role in Neurogenesis: Possible Implication for Parkinson's disease. J Exp Neurosci. 12:1179069518779829. [PMC free article] [PubMed]
Nadim, F., & Bucher, D. (2014). Neuromodulation of neurons and synapses. Current opinion in neurobiology, 29, 48-56.
Pasternak, J. F., & Woolsey, T. A. (1975). On the "selectivity" of the Golgi-Cox method. J Comp Neurol, 160(3), 307-312. doi: 10.1002/cne.901600304
Perea, G., Sur, M., & Araque, A. (2014). Neuron-glia networks: integral gear of brain function. Frontiers in cellular neuroscience, 8, 378.
Smit, G. J., & Colon, E. J. (1969). Quantitative analysis of the cerebral cortex. I. Aselectivity of the Golgi-Cox staining technique. Brain Res, 13(3), 485-510.
Sterling, P., & Laughlin, S. (2015). Principles of neural design. MIT press.
University of Queensland, Queensland Brain Institute, Types of Neurons, n.d. https://qbi.uq.edu.au/brain/brain-an.../types-neurons; Retrieved 8/31/21
Van Essen, D., & Kelly, J. (1973). Correlation of cell shape and function in the visual cortex of the cat. Nature, 241(5389), 403-405.
Attributions
Communication within the Nervous System, Neurons and Synaptic Transmission by Kenneth A. Koenigshofer, PhD, Chaffey College, is licensed under CC BY 4.0
"Types of Neurotransmitters" and "Glia Release Transmitters Too" adapted by Kenneth A. Koenigshofer from Sukel, K (2019) Neurotransmitters from https://dana.org/article/neurotransmitters/
Outside Resources, and some text adapted by Kenneth A. Koenigshofer from Furtak, S. (2021). Neurons. In R. Biswas-Diener & E. Diener (Eds), Noba textbook series: Psychology. Champaign, IL: DEF publishers. Retrieved from http://noba.to/s678why4
Neurons by Sharon Furtak is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License. Permissions beyond the scope of this license may be available in our Licensing Agreement.
Figures 1 and 2 and some text adapted by Kenneth A. Koenigshofer from: General Biology (Boundless), Chapter 35.2C, The Nervous System; Synaptic Transmission, https://bio.libretexts.org/Bookshelv...gy_(Boundless); LibreTexts content is licensed by CC BY-NC-SA 3.0. Legal.
Additional revisions made by Alan Keys, Ph.D., Sacramento City College, Sacramento, CA.