The publication of The Origin of Species by Charles Darwin in 1859 became an intellectual flash point in European intellectual life (Darwin 1859). It was focused on a significantly narrow point: Where do new species, adapted to their surroundings, come from? The Bible says God made all species. However, the Bible also says that God made all languages at the foot of the Tower of Babel; and yet, half a century of historical linguistics had showed clearly that such was not the case (French and Spanish had only been different languages, having diverged from Vulgar Latin, for a matter of a few centuries), and nobody seemed to get too upset about it.
Moreover, the suggestion that species came from other species was not all that radical. The celebrated French naturalist Lamarck had said as much in 1809 and an anonymous 1844 English bestseller called Vestiges of the Natural History of Creation had sensationalized it—to the consternation of both theologians and naturalists. Indeed, by the 1850s European biologists were very confident that cells were fundamental units of life and that the only way you could get new cells was from old cells. While this begged the question of where the first cell came from, it nevertheless was not too much of a stretch to see species as fundamental units of life as well and to ask whether new ones arose miraculously, or just from older species. The idea that species had their beginnings in other, older, similar species was known as “the transmutation hypothesis.”
Charles Darwin had come to think about the origin of species upon returning from a long voyage around the world in the early 1830s on the H.M.S. Beagle. In South America, Darwin had observed that the unusual species he saw alive there were very similar to the unusual extinct animals in the same area. This suggested some sort of historical continuity between them—descent with modification, he called it. The problem was how to make sense historically, rather than miraculously, of the particular adaptations that differentiate species. The engine of adaptation, Darwin realized, was competition. This did not necessarily entail face-to-face competition but simply the fact that not all members of a species are equally likely to survive and breed. Which ones are more likely? The ones that randomly are a bit more in sync with their environment. Those creatures will disproportionately thrive and breed, and the next generation of the species will come to look just a bit more like them, on the average. The core of Darwin’s thought is thus a two-step process: the random generation of variation, and the nonrandom process by which the environment subtly favors organisms with certain features to thrive and breed.
The biology that Darwin learned in college had invoked a famous simile: a species is like a watch, meticulously crafted by a wise watchmaker, implying a heavenly species-maker. Darwin substituted a more powerful simile, arguing that a species is actually like a breed or strain of animals, rather than like a watch. But we know that a breed or strain of animals arises naturally, historically, by the actions of breeders who select certain features to characterize populations.
Whether dogs, pigeons, or roses, the properties of living beings can change, and have changed, in quite dramatic ways by virtue of human activity in rather short periods of time. If people could make beagles and greyhounds and bulldogs by selecting the progenitors of particular stocks, then maybe nature could work to select progenitors as well, although more subtly and over vastly longer periods of time (see Figure 2.5).
Figure\(\PageIndex{1}\): A Beagle.Figure \(\PageIndex{2}\): Bulldog.Figure \(\PageIndex{3}\): Greyhound
Darwin called this principle “natural selection” and planned to write a long book about it someday. But in 1858 he received a manuscript from a fellow naturalist, Alfred Russel Wallace, who had come up with quite similar ideas to his own while working in the Malay archipelago. Darwin’s friend, the geologist Charles Lyell, had papers by Darwin and Wallace read into the record, The Transactions of the Linnaean Society, July 1, 1858, so they could share credit for the discovery, and Darwin set about to publish the work he had done on natural selection. The result was called On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life, published on November 24, 1859.
Darwin’s central thesis was that the differences among breeds or strains or varieties of animals and plants were the same kinds of differences that exist between species, only smaller and formed over short periods of time. The origin of new species lay in the long-term biases of survival and reproduction in older species. The result was a convincing naturalistic explanation for adaptation. Moreover, it finally explained the nested pattern of similarities among species that Linnaeus had discovered a century earlier but couldn’t explain. Those nested patterns were the legacy of common ancestries; they were literally family resemblances.
Darwin was especially careful to omit any discussion of people from his book. He wanted the discussion to be about the general process; consequently he wrote just a single line, near the end, about people: “Light will be thrown on the origin of man and his history” (Darwin 1859, 488). He was willing to acknowledge the possibility that life had “been originally breathed into a few forms or into one,” but he was satisfied with having described the mechanism by which adaptive change has taken place in the organic world since then—in parallel with Isaac Newton, who famously refused to speculate on where gravity came from, focusing instead only on how it works (Darwin 1859, 490).
People, however, were bound to be the central issue. A British scholar named Herbert Spencer had also come up with a similar idea, which he called “survival of the fittest” and he convinced Darwin that his phrase was synonymous with “natural selection.” And of course, who was more fit than wealthy, British white men? This confusion of human history (that is, the construction of social and political hierarchies) for evolutionary biology would prove to be a consistent irritation for students of human diversity and ancestry. Indeed, this issue eventually led Darwin and Wallace to part ways. Wallace asked: if natural selection does not produce useless organs, then why does the “savage” have a brain as big as a civilized European’s, if the savage doesn’t use it? This seeming paradox led Wallace into spiritualism and the possibility that all species of organisms had evolved…but human intelligence had had a little divine help. Darwin wrote him, “I hope you have not murdered too completely your own and my child” (Darwin, 1869). In 1871, the early British anthropologist Edward Tylor formally separated the evolution and study of “culture” from the biological properties of people. Of course the so-called “savage’s” brain was as good as the European’s, and he does use it fully, but it was filled with different information—“knowledge, belief, art, morals, law, custom, and any other capabilities and habits acquired by man as a member of society” (Tylor 1871, 1). Furthermore, this cultural information was the product of historical process, not miracle. This understanding marks the beginning of modern anthropology.
Within the academy, there was not too much reaction against the proposition that humans had descended with modification from an ape stock, and had then differentiated from that stock over the eons as a result of the differential preservation of favorable variations. The heart of Darwinism as applied to humans is simply ape ancestry and adaptive divergence.
Figure \(\PageIndex{4}\): The frontispiece to Ernst Haeckel’s (1868) popular German book on Darwinism. The English translation lacked this illustration.
But the early Darwinians were faced with a dilemma—in 1860, there was no fossil evidence linking humans to apes. The German biologist Ernst Haeckel solved this problem by fatefully arguing that we don’t need a fossil record to link us to the apes, because Europeans are linked to the apes through the nonwhite peoples of the world. He envisioned 12 different species of living peoples, each at different distances from the apes, thus sacrificing the full humanity of most people on the altar of Darwinism (see Figure 2.6). Scientists of the 1860s thought the full humanity of Africans was less important than evolution, Today that is morally repugnant. While Darwin and his English colleagues did not agree with these details, they nevertheless saw Haeckel as an ally in the broader struggle to get evolution accepted. With hindsight, we can judge this to be a morally questionable decision: Today we would hopefully universally consider the full humanity of Africans to be more important than whether humans are descended from apes, and thoroughly repudiate anyone who denied it.

 Figure \(\PageIndex{1}\): A Beagle.
Figure \(\PageIndex{1}\): A Beagle. Figure \(\PageIndex{2}\): Bulldog.
Figure \(\PageIndex{2}\): Bulldog. Figure \(\PageIndex{3}\): Greyhound
Figure \(\PageIndex{3}\): Greyhound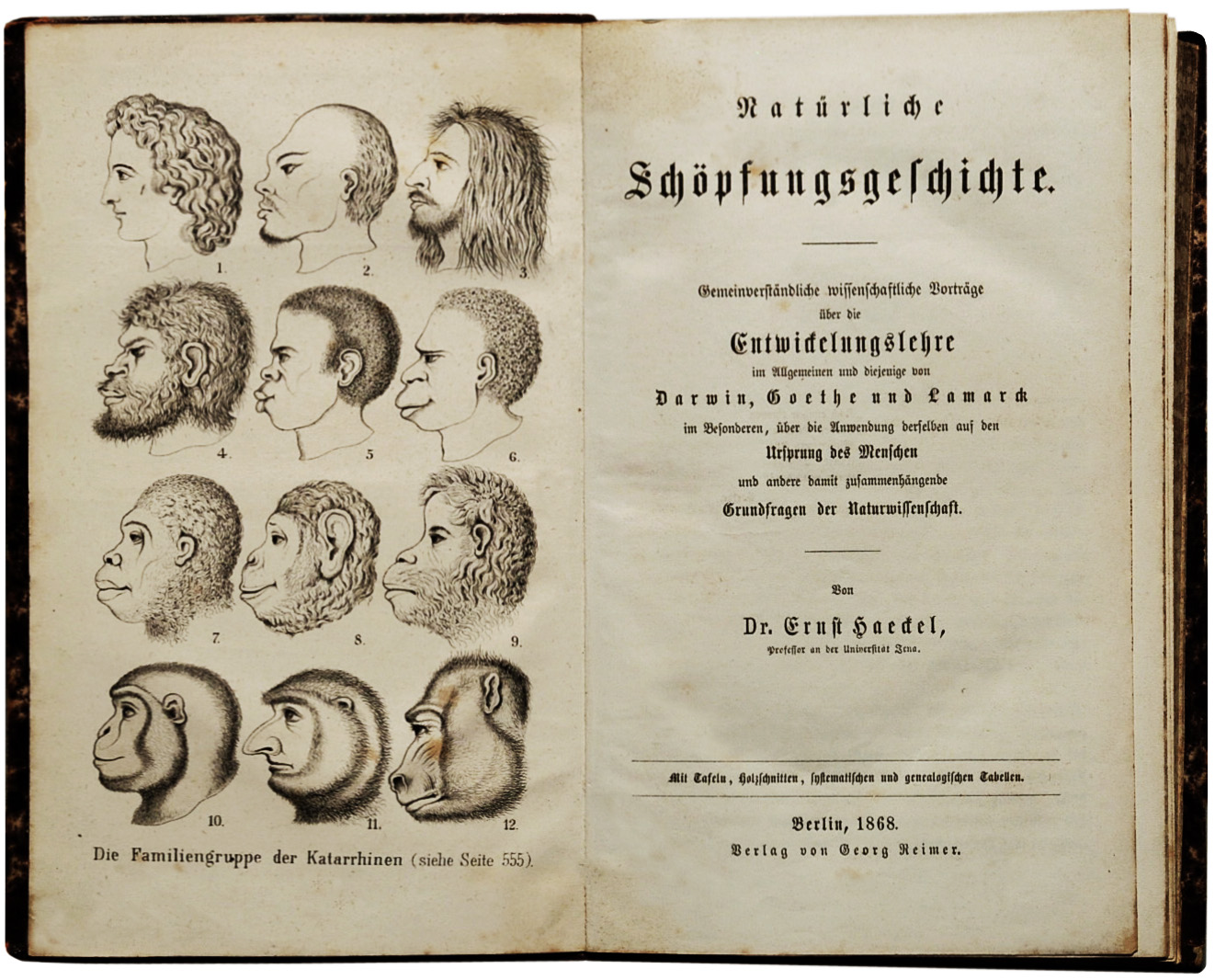 Figure \(\PageIndex{4}\): The frontispiece to Ernst Haeckel’s (1868) popular German book on Darwinism. The English translation lacked this illustration.
Figure \(\PageIndex{4}\): The frontispiece to Ernst Haeckel’s (1868) popular German book on Darwinism. The English translation lacked this illustration.