The Evolutionary Synthesis successfully reduced evolution to genetics, but until the 1980s it was not possible to study the DNA sequence of the genes directly. Various surrogate measures had been employed for decades. For example, not only is blood a powerful metaphor for heredity, but also it contains genetically controlled immunological properties that can be used to study evolution. It was known in this way by the 1920s that the blood of human and chimpanzee were more similar to one another than were the blood of horse and donkey (see Figure 2.9). By the mid-1960s, it was well established that the blood of human and chimpanzee were more similar to one another than either was to the blood of an orangutan. With greater precision, the actual amino acid sequences of some proteins could be established and compared across species.
It quickly became clear that while genetic differences appear generally to track anatomical differences—that is, the closest relatives of species inferred from their hemoglobin (the blood protein that carries gases) are generally the same as those inferred from their teeth—they nevertheless don’t match well quantitatively. Thus, while humans are very easily distinguishable from gorillas physically and mentally, their hemoglobins only have two differences—the other 285 amino acids composing the protein match up perfectly. With less than one percent difference in the structure of their hemoglobin, yet striking differences in anatomical form, communication, and behavior, there seems to be a paradox in their biochemical versus anatomical relationships. This led to some thoughtless early inferences from biochemists, such as suggesting that humans are merely variant gorillas, from the viewpoint of hemoglobin. (But if we do not appear to be variant gorillas from any other viewpoint, then perhaps the viewpoint of hemoglobin—or molecular genetics more broadly, so went the counter-argument—is a foolish one to adopt.)
Figure \(\PageIndex{3}\): Dentition of Ramapithecus.
We now appreciate that anatomical variation tracks adaptive divergence of the species (obvious differences between humans and apes relate to locomotion, cognition, sound production, heat dissipation, etc.). But genetic variation more closely tracks the time since the species diverged from one another. By the late 1960s, molecular data were being used to test an important hypothesis about human evolution. Where physical anthropologist Sherwood Washburn thought that humans and African apes probably shared a common ancestor as recently as three to five million years ago, paleoanthropologist David Pilbeam felt that they had separated far earlier than that. Armed with the well-dated (but poorly reconstructed) dental remains of a 14-million-year-old fossil called Ramapithecus, Pilbeam argued that Ramapithecus was a part of the human lineage, which in turn had to be at least that old (Figure 2.10). But Washburn’s colleagues, Allan Wilson and Vincent Sarich, showed in 1967 that (1) the biochemical changes they measured were changing in a clocklike manner and (2) given the small amount of biochemical difference detectable between human and chimpanzee, the species separated no more than five million years ago. Thus, (3) Ramapithecus could not be on the human line 14 million years ago, because there was no separate human line 14 million years ago!
We now see Ramapithecus differently, as part of the orangutan lineage, and we find that genetic or molecular evolution does indeed tend to track time, rather than adaptive divergence. The reason is that most of the genome’s DNA falls between genes and does not actually code for anything. Consequently, mutations that occur to most of the DNA do not have discernible effects on the body and are thus nonadaptive. Only a small bit of the DNA, it seems, actually builds the organism and encodes its adaptations; and even today, the processes by which it does so are vaguely understood.
When we compare actual DNA sequences across species, we consequently find striking patterns. Notably, we almost always find more difference across species in DNA between genes than in DNA within genes (see Figure 2.11). Where you might find two percent difference between species in the base sequence of a gene, you will find three percent difference in the DNA outside of that gene. Mutations are just as likely to arise within a gene as outside of a gene, yet when you compare species, you find more differences between genes. This suggests that the DNA between genes can tolerate changes without significantly harming the organism, because that DNA is not expressed, while DNA within genes cannot tolerate mutations quite as readily, so they get weeded out. Why? Because the genes do indeed function; consequently, random changes in a gene are far more likely to compromise that function than to improve it. Imagine trying to adjust the fuel injector in your car with a hammer. There is a small probability that you might hit it in just the right way to improve its performance, but chances are good that you would make it worse. Similarly, a random change to an already-functioning molecule is far more likely to make it work worse than to make it work better. That is why mutations can give you cancer, not superpowers. And by compromising the health of its bearer, such a mutation would be “weeded out” by natural selection (See the discussion in Chapters 3 and 4).
Figure \(\PageIndex{4}\): DNA comparisons yield more difference between than within genes.
This interpretation is supported when we examine the DNA differences simply within genes across species. While most mutations to the gene’s coding sequence must affect the structure of the protein it codes for, a few do not. We call these “synonymous mutations,” and when we compare genes across species, we almost always find far more of them than we find of the mutations that do indeed change the structure of the gene product. So even though synonymous mutations are a small proportion of mutations, they predominate in cross-species comparisons of genes. And for exactly the same reason: synonymous DNA mutations are less likely to be weeded out, because they are unexpressed and are thus invisible to the environment.
This helps to explain why the genetics seems to track time while the anatomy seems to track adaptation. If most mutations are neutral, with no net effect on the fitness of the organisms that possess them, then (as statisticians calculated in the 1960s) they will spread through a population rarely and in proportion to the rate at which they arise. The mutation rate is a constant, so consequently, over time, neutral mutations will spread and come to differentiate populations in proportion to the time since those gene pools have been separated from one another. Bodily difference, by contrast, interacts with the environment in important ways, and its evolution will track that interaction. Thus, biologists often envision evolution working on different hierarchical “levels”: a genetic or molecular level and an anatomical level.
Yet how do we simultaneously accommodate the knowledge that (1) genetics and anatomy are different levels, with one tracking time and the other adaptive divergence, and that (2) the genes somehow cause the anatomy? The disconnect lies in the recognition that we still do not know how our one-dimensional DNA nucleotide sequence encodes a four-dimensional animal. This was the unfulfilled promise of the Human Genome Project in the 1990s: This Project produced the complete DNA sequence of a human cell in the hopes that it would reveal how human bodies are built and how to cure them when they are built poorly; however, that information has remained elusive. Presumably the knowledge of how organisms are produced from DNA sequences will one day permit us to reconcile the discrepancies between the patterns we see in anatomical and molecular evolution.

 Figure \(\PageIndex{1}\): Horse.
Figure \(\PageIndex{1}\): Horse. Figure \(\PageIndex{2}\): Donkey.
Figure \(\PageIndex{2}\): Donkey. Figure \(\PageIndex{3}\): Dentition of Ramapithecus.
Figure \(\PageIndex{3}\): Dentition of Ramapithecus.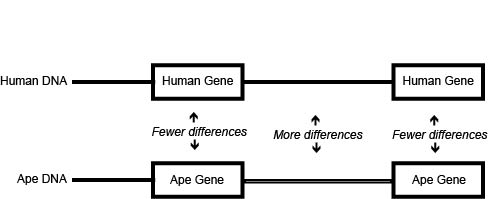 Figure \(\PageIndex{4}\): DNA comparisons yield more difference between than within genes.
Figure \(\PageIndex{4}\): DNA comparisons yield more difference between than within genes.