
14.2: Cognition, Intelligence, and Language as Psychological Adaptations
- Page ID
- 113218
This page is a draft and under active development. Please forward any questions, comments, and/or feedback to the ASCCC OERI (oeri@asccc.org).
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)- Explain what is meant by the claim that psychological processes, including intelligence, serve adaptive movement
- Explain why intelligence and cognition are examples of psychological adaptations
- Explain why natural selection requires recurrent, across-generation conditions to generate adaptations
- Describe some of the environmental regularities, including several universal recurrent relational features of the world, that may have been genetically incorporated into brain organization generating intelligence, including general "fluid" intelligence
- Discuss Spearman's definition of "g"
- Describe the brain areas that are involved in intelligence, broadly defined as brain systems that guide behavior toward successful adaptation
- Discuss the role of Shepard's concept of "genetic internalization" of biologically important environmental regularities in the evolution of intelligence
- Discuss three abstract relational regularities of the world which may have been genetically internalized into the brain operations of general intelligence
- Describe vector coding
- Discuss mental models, prospection, and intelligence, and list brain structures most directly involved in the formation of mental models/cognitive maps
- Describe the neural correlates of differences in intelligence across species and the P-FIT theory of differences in intelligence among humans
- Describe the Broad Visual Perception Factor and the role it plays in the intelligent control of movement/behavior
Intelligence, Cognition, Language, and Biological Adaptation
by Kenneth A. Koenigshofer, PhD.
Overview
After a general review of cortical functions in the previous section, we now examine higher cognitive functions as modes of achieving biological adaptation to the environment. In a previous chapter, you learned about the neural mechanisms of movement. Let's start this module with the premise that, to be useful, movement must be effectively guided in order for it to be responsive to environmental demands and adaptive opportunities. The senses are the first tier in the systems that guide movement (even some single-celled organisms can sense harmful chemicals in their fluid environment and then use their cilia to swim away from them, showing how even a simple guidance system for movement improves adaptation). Imagine you were one of our evolutionary ancestors; if you couldn't sense an approaching saber-toothed tiger and make accurate judgments about its distance and speed, it is obvious that you would be unable to effectively run or hide from it. If you couldn't feel fear and understand danger, you would not be motivated to run or hide. Without senses and perception, it would be impossible for you to organize movements to find food or secure a mate.
However, in addition to the senses, perception, and emotion as guidance systems for movement, higher order functions of the brain, some of which were discussed in the chapter on the mechanisms of movement (e.g. premotor cortex, parietal cortex), are also critically involved in the more advanced control of action. In this chapter, we consider intelligence and cognition as part of the sophisticated systems that guide movement in animals and humans to maximize their adaptation to the environment.
We will also consider other research on intelligence with a very different focus. Early work by psychologists interested in human intelligence took a more practical approach. These psychologists in the first part of the twentieth century focused on the development of methods of measuring intelligence in humans. This approach to the study of psychological attributes is called psychometrics. Though early research emphasized the creation and refinement of intelligence tests, rather than the biological origins and functions of intelligence, psychometric analysis of the performance of large groups of people on intelligence tests (Spearman, 1904, 1925) ultimately led to theories of the structure of human intelligence (Carroll, 1993; Cattell, 1987). These theories are important to biological psychology because they have influenced much of the modern thinking and research on the genetic and brain mechanisms involved in human thinking and intelligence. Additional details about psychometrics and theories of intelligence can be found in Supplement 1, Traditional Models of Human Intelligence, in this chapter.
More recently, intelligence research by psychologists and neuroscientists has expanded to include the study of brain mechanisms underlying intelligence in humans and in other species. This approach has led to conceptions of intelligence and cognition in a broader biological and evolutionary context. On this view, thinking, intelligence, and language are products of evolution, just like other genetic traits of organisms. Thinking, intelligence, and language exist because they helped ancestral humans meet the challenges presented by the environment, challenges which drove natural selection. In short, thinking, intelligence, and language are biological in origin, biological in function, and, of course, they are generated in a biological organ, the brain.
One of the key evolutionary trends responsible for the evolution of intelligence and cognition was genetic incorporation of information about biologically significant regularities of the world into the circuitry and operations of the brain. As a consequence, the brain is equipped with many "cognitive instincts" and innate implicit knowledge about biologically important, enduring regularities of the terrestrial environment, forming the groundwork for much of our genetically evolved intelligence as a species.
This view contrasts with and rejects the "blank slate" view of the mind/brain assumed in the Standard Social Science Model (SSSM), the set of assumptions that much of psychology and the social sciences were founded upon--the view that humans lacked any innate psychological nature and that the mind and brain were essentially blank at birth, leaving it to learning and culture to form human behavior free of genetics and biological evolution. In this chapter, we take a different point of view--that biology, genes and evolution, are the primary determinants of human cognition and behavior, in interaction with learning and cultural influences, which themselves are ultimately biological in nature.

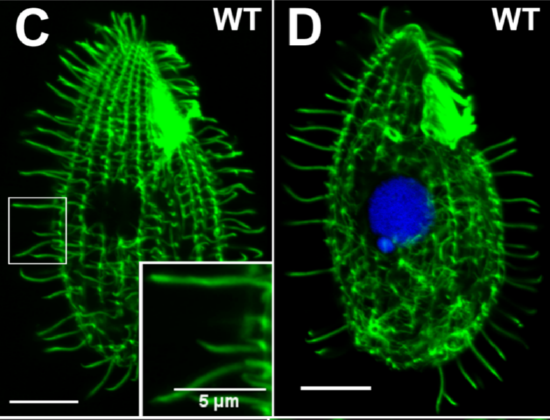
Figure \(\PageIndex{1}\): (Left) Lioness hunting in the Serengeti region of Tanzania. Its mechanisms of sensation and perception, emotion, and intelligence are pitted against these same guidance systems for movement found in its prey. Predator-prey interactions may have been an escalating evolutionary impetus for the development of intelligence in both predator and prey. (Right) A single-celled organism featuring its cilia which it uses to move toward favorable parts of its fluid environment and away from harmful regions, thereby facilitating its adaptation. Approach to beneficial elements of the environment and avoidance of and withdrawal from harmful elements is a primary rule governing guidance systems for movement in all motile species. (Image on left is from Wikimedia Commons, https://commons.wikimedia.org/wiki/F..._saturated.jpg, by Schuyler Shepherd; licensed under the Creative Commons Attribution-Share Alike 2.5 Generic license. Caption by Kenneth A. Koenigshofer, PhD. Image on right is from Wikimedia Commons, https://commons.wikimedia.org/wiki/F...C3%BAcleos.png, by Seixas C, Cruto T, Tavares A, Gaertig J, Soares H; licensed under the Creative Commons Attribution 4.0 International license. Caption by Kenneth A. Koenigshofer, PhD).
Thinking and intelligence serve the adaptive organization of movement
As argued in the chapter on evolution and genetics, nothing makes sense in psychology except in the light of evolution. This claim is especially relevant for a course in biological psychology. When we consider thinking and intelligence from a biological and evolutionary perspective, it is important to ask what functions they perform for the organism. The brain does all kinds of complex processing, but it is important to understand that for that processing to have any effect on the environment, brain activity must ultimately converge onto motor neurons in the spinal cord ("the final common path") that stimulate the muscles to produce movement--behavior (see the chapter on movement). Like every aspect of our psychology and its corresponding brain activity, the function of thinking and intelligence is to generate adaptive behavior, movement, to successfully meet environmental challenges to survival and reproduction and to exploit opportunities, thereby increasing biological fitness.
Consider plants, for a moment. They don't have intelligence or thinking--they don't need to, because they depend very little upon movement for survival and reproduction--instead, water and their source of energy, sunlight, come to them (plants can move slowly in a limited way; they grow toward sunlight and roots grow toward water sources, but contrast this with all the complex social behaviors modern humans engage in to get food and water--e.g. agriculture, supply chains, plumbing, dams, water companies, etc.). Neither do plants have to flee or hide from predators; they protect themselves with thorns or poisons or simply grow back if partially eaten. Neither do they need to move for reproduction--wind and insects and other animals carry the reproductive cells for them (imagine if this were true for humans! What would happen to human courtship, dating, and romantic love? The purpose of which, from a biological perspective, is to get egg and sperm cells together).
But none of this is true for animals. For animals, including the human animal, movement is key to survival and reproduction, and, as noted above, the movement must be guided to form patterns of action that solve adaptive problems and exploit adaptive opportunities. According to Darwin's theory of evolution, which he explained as "descent with modification," the roots of human cognition and intelligence lie deep in our species' evolutionary ancestry. Evidence for this can be found in Darwin's principle of the "continuity of species" applied to psychology, as shown by the great degrees of

Figure \(\PageIndex{2}\): Movement is essential for survival and reproduction in animals. The great Wildebeest migration with over 2 million animals follows the rains to lush new feeding grounds. (Image from Wikimedia Commons; https://commons.wikimedia.org/wiki/F...n_crossing.jpg by Naturaltracksafaris; licensed under the Creative Commons Attribution-Share Alike 4.0 International license. Caption by Kenneth A. Koenigshofer, PhD).
similarity in the guidance and control systems for movement and their underlying brain mechanisms across mammalian species--for example, all mammals have similar brain structures, including limbic structures and cerebral cortex. All species respond to potentially harmful stimuli with threat, attack, or escape. All mammals respond to sexual signals by moving toward and making contact with their source. Each suggests common behavioral control mechanisms evolutionarily conserved across species
But intelligence also exists in non-mammalian species. Intelligence in distantly related species including some birds, such as Corvids (e.g. ravens, jays, crows) and parrots, and in invertebrates such as the octopus and cuttlefish (Adams & Burbeck, 2012; Mather, 2019; Mather & Dickel, 2017), illustrate convergent evolution, suggesting the high adaptive utility of intelligence in diverse ecological niches. Intelligence and thinking, like all psychological processes, evolved to serve the organization of movement to make behavior adaptive, successful, in the Darwinian struggle for survival and reproduction (Koenigshofer, 2011, 2016).
Cognition and Intelligence are psychological adaptations
Intelligence and thinking (i.e. cognition) are psychological adaptations evolved by natural selection over millions of years. A psychological adaptation is a psychological or behavioral trait that has developed through evolutionary processes such as natural and sexual selection and which is encoded into a species' DNA (see Ellis & Ketelaar, 2002). Dicke and Roth (2016) offer a comprehensive definition of intelligence recognizing its role in adaptation to the environment in a wide range of animals and humans:
According to the majority of behaviorists and animal psychologists, ‘intelligence’ can be understood as mental or behavioral flexibility or the ability
of an organism to solve problems occurring in its natural and social environment, culminating in the appearance of novel solutions that are not
part of the animal’s normal repertoire. This includes forms of associative learning and memory formation, behavioral flexibility and innovation rate, as well as abilities requiring abstract thinking, concept formation and insight.
By contrast, Colom et al. (2010) define intelligence simply as "a general mental ability for reasoning, problem solving, and learning."

Figure \(\PageIndex{3}\): Because animals cannot photosynthesize, they must move in order to obtain their sources of energy found in plants and other animals--animals which can hide, run, and fight back--unlike the plants' source of energy, the sun. Movement is organized by brain circuitry which has been configured by natural selection over evolutionary time to solve adaptive problems. Here a hyena on the plains of Africa solves the problem of getting sufficient energy to survive and ultimately reproduce its genes. (Image from Wikimedia Commons; https://commons.wikimedia.org/wiki/F...il_breghys.jpg; by Brwynog; licensed under the Creative Commons Attribution-Share Alike 4.0 International license).
A typical definition of general intelligence (discussed below) shows how intelligence and general intelligence are terms sometimes used interchangeably, even by psychologists. General intelligence is often defined as "ability to reason deductively and inductively, to think abstractly, use analogies, to synthesize information, and to apply that information to new domains" (Gottfredson, 1997; Neisser et al., 1996).
In spite of some differences in definition among researchers, there is unanimous agreement that intelligence and cognition are properties of brain processes which have been shaped and refined over millions of years of evolution. Once again, Darwin's principle of the "continuity of species" leads us to suspect that there are many similarities between intelligence and cognition in humans and these same processes in animals, especially mammals. To help us understand human intelligence and cognition and their evolutionary origins, many biological psychologists study these processes in other animals, including apes, monkeys, rats, dogs, parrots, and Corvids, such as ravens, jays, and crows. Animals often surprise us with the sophistication of their emotional sensitivity and their intelligence even though they lack language (Huber & Gajdon, 2006; Wasserman, et al., 2006). An interesting anatomical feature of the cerebral cortex is what we might call microprocessors or "chips" in the human cortex. These are columnar structures, known as cortical columns, containing approximately 3,000 neurons each. There are approximately 150,000 cortical columns in the human cerebral cortex (Hawkins, et al., 2017) . These structures are very similar from one part of the cortex to another and from one mammalian species to the next and suggest highly generalized computational functions in cerebral cortex, whereas specialized modules or genetically dedicated circuits for processing of emotional and motivational information are localized to subcortical regions of the brain, shared by all mammals (Panksepp & Panksepp, 2000).
Intelligence and cognition (i.e. thinking) are exceedingly complex processes. As a consequence, the brain mechanisms involved in these processes are not well understood. As you can see, even definitions of these processes vary widely in the scientific community. However, all of the definitions above include terms such as reasoning, abstract thinking, insight, or problem solving, processes which themselves are not well defined or clearly understood by psychologists and neuroscientists. This highlights the fact that psychologists and neuroscientists are still in the early stages of gaining a real understanding of how intelligence and thinking work and what neural processes in the brain produce them. Perhaps this is why textbooks in biological psychology have tended to give little or no attention to these processes even though they are perhaps the most wondrous and important products of brain activity.
Psychometrics, the Measurement of Human Intelligence, and "g"
Early in the history of psychology, research about intelligence was not focused upon its evolutionary origins or its adaptive functions. Instead, psychologists concentrated their research efforts on more practical problems such as the measurement of intelligence for classifying students and military recruits. Major study of human intelligence by psychologists began in the early 1900's when psychologists focused on the measurement of intellectual abilities, an approach known as psychometrics. This research approach resulted in the development of intelligence tests and the concept of IQ (intelligence quotient). Additional research in psychometrics, focused on mathematical analysis of performance scores of large populations on IQ tests using a method called factor analysis, which analyzes patterns of correlations between measures. This research led to the finding that people who did well on any particular measure of intellectual performance tended to do well on all such measures. This led psychologist, Charles Spearman (1904), to hypothesize the existence of a single, unitary "g" factor (g for general) in human intelligence. This g factor referred to a general mental capacity which was hypothesized to be involved to varying degrees in multiple mental abilities including verbal, spatial, mechanical, computational, and other performances. "This g factor provides an index of the level of difficulty that an individual can handle in performing induction, reasoning, visualization, or language comprehension tests" and the g factor accounts for "more than half" of the variation among individuals "in a cognitive test or task" (Jung & Haier, 2007, p. 492). The g factor came to be known among psychologists as "general intelligence." Spearman believed that "g" was most closely related to what he called the "eduction [from the Latin root "educere" which means to "draw out"] of relations and correlates," important in inductive and deductive logic, grasping relationships, inferring rules, and recognizing differences and similarities. In the sections below, we examine some key components of intelligence, relevant brain mechanisms, and how intelligence might have evolved.
Conditions required for evolution of psychological adaptations
Understanding the evolutionary origins of intelligence and thinking may offer insights about how they work and about their underlying brain mechanisms. First of all, remember the claim made above that intelligence and cognition are psychological adaptations, or more correctly, a collection of psychological adaptations, each evolved by natural selection to process certain types of information in specific ways. A key principle of how evolution works is relevant here. Evolution by natural selection can only capture information about features of the world that are present generation after generation. This means that any innate brain mechanisms and the innate "instincts" produced by these mechanisms must have been formed as adaptations to regularly recurrent properties of the environment. This is because natural selection requires many generations to work a genetic change in a population (see chapter on evolution and genetics). Without consistent (statistically regular) selection criteria present over many generations, natural selection cannot fashion complex adaptations. For example, if the consistent force of gravity had not been regularly present over generations, land-living vertebrates could not have evolved bones with tensile strength sufficient to adapt to the downward pull of gravity. In this case, these animals would be incapable of movement or even standing. Another example is color vision. The color of many fruits indicates their state of ripeness. Without the ripeness of fruit being regularly signaled by its color, generation after generation, color vision would not have evolved in fruit-eating primates. Stated another way, information about the strength of gravity or about the correlation between wavelength of light and ripeness of fruit could not have been incorporated by natural selection into the evolved adaptations of organisms unless these features of the world were consistently present, and affecting rates of survival and reproduction, generation after generation.
By this reasoning, long-term, across-generation regularities of the world that have adaptive significance should be expected to play a special role in the evolution of the mind/brain (just like they do in the evolution of anatomical and physiological adaptations). As evolutionary psychologists, Tooby and Cosmides (1992, p. 69), state: “Long-term, across-generation recurrence of conditions ... is central to the evolution of adaptations." Kaufman et al. (2011, p.213) express a similar idea. They state, "Evolutionary psychologists sometimes argue that a class of situations must be relatively narrow to exert consistent selection pressure, but this claim is insufficiently justified. Any regularity in the environment can exert selection pressure if it poses a challenge or an opportunity to the organism . . ." And as former Stanford psychologist, Roger Shepard, put it, there has evolved "a mesh between the principles of the mind and the regularities of the world" (Shepard, 1987a). This means that the organization of the mind reflects regularities of the world, because of the fact that natural selection operates on conditions that repeat, that are regular, over long periods of time. So, when thinking about the evolution of brain and mind, we should be on the lookout for "instincts" that innately predispose the brain to process environmental inputs using innate principles derived from adaptively significant regularities of the world. In other words, the brain is filled with circuitry that operates by genetically programmed rules derived from information about enduring environmental regularities.
Examples of such long-term regularities or enduring properties of the world are: the widespread presence of environmental stimuli that can damage body tissue; that some things in the environment contain sources of nutrition while others are poisonous; that some potential mates are more likely than others to be healthy and fertile; that snakes and spiders are frequently dangerous; that sugars and fats are concentrated sources of energy; that high status in the social group in social animals gives greater access to resources including mates; and so on. In response to biologically important statistical regularities in the world such as these, neural circuitry has evolved in animal and human brains that causes: withdrawal from harmful stimuli in response to pain; approach toward and consumption of sources of nourishment and avoidance of potential foods that cause illness; powerful innate drives and emotions to mate with sexual partners possessing features indicative of health and reproductive potential (Ellis & Ketelaar, 2002, p. 162; Gangestad & Simpson, 2000); a genetic readiness in humans and other primates to learn fear of snakes and spiders (DeLoache & LoBue, 2009); a startle response to sudden loud noises; status seeking and sensitivity to social standing in social primates; a preference in humans for sugars, salt, and fats; and so on (see Tooby & Cosmides, 2015).


Figure \(\PageIndex{4}\): (Left) For most people, spiders like this one induce a strong aversive reaction motivating avoidance and withdrawal behavior, an example of a psychological adaptation evolved to protect us from possible poisoning from this class of stimulus. (Right) Humans have evolved a preference for fats, sweets, and salt, a psychological adaptation from the Pleistocene which can be harmful to health if followed too frequently in the modern urban environment full of fast food restaurants--a very different environment from that of our hunter-gatherer ancestors. (Image on left from Wikimedia Commons, https://commons.wikimedia.org/wiki/F...ia,_Brazil.jpg; by Alex Popovkin, Bahia, Brazil; licensed under the Creative Commons Attribution 2.0 Generic license. Image on right from Wikimedia Commons, https://commons.wikimedia.org/wiki/F..._Bigntasty.jpg; by مانفی in Persian; licensed under the Creative Commons Attribution-Share Alike 4.0 International license. Captions by Kenneth A. Koenigshofer, PhD).
Each of these innate behaviors evolved in response to specific regularities of the world, "recurrence of conditions," that consistently had important adaptive consequences--consequences for survival and reproduction--over countless generations of evolution.
.jpg?revision=1&size=bestfit&width=508&height=460)
Figure \(\PageIndex{5}\): According to evolutionary psychologists, Cosmides and Tooby (2003), forming and maintaining friendships is important to us today because this was one of many adaptive problems that our human ancestors encountered and had to solve in our Pleistocene past. Friendships led to alliances that were important in securing resources within Pleistocene bands of individuals that depended upon one another for survival. For this reason, we are motivated to find friends and having them feels good, reinforcing the behavior. Information processing by the brain necessary for formation and maintenance of relationships is one of the topics studied by biological psychologists interested in intelligence and social cognitive neuroscience. (Image from Wikimedia Commons, https://commons.wikimedia.org/wiki/F...923360765).jpg; by Rod Waddington; licensed under the Creative Commons Attribution-Share Alike 2.0 Generic license. Caption by Kenneth A. Koenigshofer, PhD.).
Evolutionary psychologists have identified some additional recurrent problems that had to be regularly solved by our human ancestors: " . . . winning social support from [group] members, remembering the locations of edible plants, hitting game animals with projectiles, …, recognizing emotional expressions, protecting family members, maintaining mating relationships, …, assessing the character [and social valuations] of self and others, causing impregnation, acquiring language, maintaining friendships, thwarting antagonists, and so on" (Cosmides and Tooby 2003, p. 59).
In each case, evolutionary psychologists suspect that brain mechanisms have evolved to contribute to solutions of each of these adaptive problems regularly present generation after generation. On this view, cognition is believed to consist of “many mental rules that are specialized for reasoning about various evolutionarily important domains, such as cooperation, aggressive threat, parenting, disease avoidance, predator avoidance, object permanence, and object movement” (Cosmides & Tooby, 1992, p. 179). But these are not the only kinds of recurrent, across-generation conditions that have been genetically represented by natural selection in brain circuitry. Such regularities can be much more abstract and widely distributed throughout the environment.
Innate (instinctual) Knowledge of Universal Regularities of the World: The Foundations of Intelligence
One approach to understanding the nature and origins of intelligence is the idea that natural selection incorporated information about “general—perhaps even universal—properties” of the world (Shepard, 1992, p. 500) into brain organization over evolutionary time (Shepard, 1992, 1994, 2001). The core idea is that natural selection has selected genes that equip human and animal brains with fundamental information about how the physical world is organized, information that is essential for intelligent adaptation to the environment. This inborn information provides innate "knowledge" about biologically significant, universal properties of the world which have existed continuously for countless generations. This approach also may explain fundamental principles of how our thinking processes are organized and how they evolved.
According to Shepard (1987a, 1992, 1994, 2001), natural selection has favored the genetic incorporation or “internalization” of biologically significant regularities of the world, “whether . . . within a particular species' local niche or throughout all habitable environments. . . . Genes that have internalized these pervasive and enduring facts about the world should ultimately prevail over genes that leave it to each individual to acquire such facts by trial and possibly fatal error” (Shepard, 1994, p.2).
What Shepard means by "genetic or evolutionary internalization" is that natural selection has favored genes that incorporate information into the brain about specific, biologically important, universal regularities of the terrestrial environment. Consequently, information about these enduring environmental regularities, such as the fact that the sun rises and sets approximately every 24 hours, becomes genetically encoded into genes which organize specific brain circuitry or other properties of the brain. In the case of the roughly 24 hour cycle of light and dark, we see daily rhythms of sleep and wakefulness controlled by inborn brain mechanisms and by cycles of melatonin release from the pineal gland, a small gland near the center of the brain, which acts on the hypothalamus, involved in the control of these daily cycles. We also see daily cycles of a number of other hormones as well. The release of these hormones is regulated by the pituitary gland which itself is under the control of genetically organized circuitry within the suprachiasmatic nucleus of the hypothalamus. Animals and humans have many so-called circadian (about a day) rhythms. These rhythms have originated from "genetic or evolutionary internalization" of information about the repeating daily cycle of light and dark into brain organization as a result of natural selection (see chapter on sleep).

Figure \(\PageIndex{6}\): A woman sleeping. She does not know that daily cycles of sleep and wakefulness and other circadian rhythms are the result of genetic internalization over the course of evolution of an enduring regularity of the physical environment, the rotation of Earth on its axis (see text below and the chapter on sleep). (Image from Wikimedia Commons; https://commons.wikimedia.org/wiki/F...g_in_Sepia.jpg; by Massimo Danieli; licensed under the Creative Commons Attribution-Share Alike 2.0 Generic license. Caption by Kenneth A. Koenigshofer, PhD.).
Some of these enduring properties of the environment which have been "genetically internalized" by natural selection (Shepard, 1992, 1994, 2001) into the brain's functioning are relatively concrete--such as the approximately 24 hour cycle of light and dark (mentioned above). Others are “three-dimensional, locally Euclidian space” with a "gravitationally conferred unique upward direction," and one-dimensional time with a "thermodynamically conferred unique forward direction" (Shepard, 1992, p. 500). As noted above, circadian rhythms built into us and other species are the result of this "genetic internalization" of the daily light/dark cycle which characterizes the terrestrial environment in all but the polar regions of Earth. The genetic internalization of information about three-dimensional space is evidenced by the presence of "place cells" and spatial cognitive maps in the hippocampus, as we discussed earlier in Chapter 10 on learning and memory, and involvement of the parietal cortex in spatial processing. And one-dimensional time has been internalized into the functional organization of the cerebellum where cells which monitor time have been identified (Irvy, et al., 1989; Irvy & Spencer, 2004; Hayashi, et al., 2014). Some additional regularities of the world that have been genetically internalized by evolution may help solidify the point. Objects in the universe tend to have certain shapes, reflectance, and paths of motion and these statistical regularities of the visual world have been "genetically internalized" (Shepard, 1992) by natural selection into the human visual system and the visual systems of many other species (Anderson, 2009). Other biologically significant regularities of the world, including regularities of the social environment, leading to evolution of other quite specific components of intelligence (including social intelligence; see sections 18.11 and 18.14) are the recurrent problems of survival and reproduction mentioned above. All of these are examples of how the principles of the mind reflect some of the more concrete regularities of the world (recall the quote above from Shepard).
Natural Selection Favored Representations of Abstract Relational Regularities: Evolution of General Intelligence
In addition to the above relatively concrete regularities of the physical world, there are other enduring and pervasive regularities of the physical environment that are quite abstract and relational and information about these have also been "genetically internalized" into brain operations by natural selection. These relational regularities of the world are cause-effect, similarity, and predictive relations between objects and events. These relations are so ubiquitous and so familiar to us that often we barely notice them. Yet they are essential to our intelligence. Natural selection has also "genetically internalized" information about these adaptively important relational regularities of the world into our brain organization. As a consequence, our brains are innately predisposed to readily understand cause-effect, to recognize similarity relations which we use to form categories and concepts, and to look for and to find predictive relations among events in the world (Koenigshofer, 2017). This means that we are born with many genetically encoded "instincts" about the abstract, yet fundamental properties of the physical environment. In effect, we inherit some general, universal principles about how the world works. These "cognitive instincts" comprise much of our genetically inherited "general" intelligence ("g," see discussion of "g" above, and also see section 18.11) as a species--properties of the brain derived from eons of evolution by natural selection (Koenigshofer, 2017). These instincts are other ways in which "there has evolved a mesh between the principles of the mind and the regularities of the world" (Shepard, 1987a). Intelligent action depends upon comprehension of causality, similarity, and predictive relations in one's environment.
Causality
Because the world is governed by natural causal laws, cause-effect relations between things regularly occur everywhere in the environment and have consistently existed since the beginning of evolutionary time. Such regularities, though abstract and relational, can drive natural selection (Koenigshofer, 2017; see Cosmides & Tooby, 1992, p. 48; Kaufman et al., 2011, p. 213). As a result, this relational property of the world has been genetically internalized, just like information about three dimensional space and the cycles of light and dark, so that our brains (and the brains of many other species) are innately "tuned" to look for and to understand cause-effect in the environment. Penn and Povinelli (2007, p. 98) state: "Animals of all taxa have evolved cognitive mechanisms for taking advantage of causal regularities in the physical world." Evidence for an innate understanding of causal relations in humans comes from experiments showing that children, from a very early age, readily understand cause-effect and use statistical regularities to tease out causes of specific events (see Gopnik, 2010, 2012; Penn & Povinelli, 2007). The inborn predisposition to understand cause-effect as a general property of the world is a central component of human and animal general intelligence and cognition (Koenigshofer, 2017). The famous 18th century British philosopher, David Hume (1748/1988), recognized this fundamental property of thought when he wrote: "All reasonings concerning matter of fact seem to be founded on the relation of Cause and Effect." A modern definition expresses the same idea: "causal reasoning [is] the ability to identify the functional relationship between a cause and its effect (Amodio, 2019).
From the standpoint of biological adaptation, implicit knowledge about causality allows intelligent creatures to navigate and exploit an enormous variety of complex causal relations, to make causal inferences and predictions, which benefit adaptation to the problems and opportunities presented by the environment. A chimpanzee who understands that putting a long stick into a termite hill will cause termites to bite onto the stick gets a lot more juicy termites to eat than a chimpanzee who does not understand this causal relationship (Goodall, 2000). Birds have been observed to toss small pieces of food that float on the surface of a pond to lure fish to the surface where they can be captured more easily. Human understanding of complex causes led to the control of fire, the invention of cooking and agriculture, the construction of shelters and tools, and other innovations making it possible for our species to successfully occupy every region of the planet.

Figure \(\PageIndex{7}\): Bonobo chimpanzee termite fishing. (Image from Wikimedia Commons; https://commons.wikimedia.org/wiki/F...r_termites.jpg; by Mike Richey; licensed under the Creative Commons Attribution-Share Alike 3.0 Unported license. Caption by Kenneth A. Koenigshofer, PhD.).
If humans and at least some animals are born with an innate disposition to understand cause and effect, we should expect to find brain structures that seem to be especially involved in perception of and reasoning about cause-effect. Brain imaging studies show that "specific brain networks are involved in the extraction of causal structure from the world" (Fugelsang, et al., 2005, p.45). Cues humans use to determine if two events are causally related include covariation (things occur together), temporal order, contiguity (closeness) in time and space, information about possible causal mechanism in the specific instance of causality being examined, and similarity between cause and effect. Analysis of immediate visual perception of causality in humans using fMRI implicates several brain structures including right middle frontal gyrus, right inferior parietal lobule, right prefrontal, right parietal, and right temporal lobes (Fugelsang, et al., 2005). Fugelsang, et al. (2005) also reported that right parietal cortex was involved in spatial cues of causality (spatial contiguity) and right temporal cortex was involved in processing temporal cues (temporal contiguity) of causation. Right prefrontal cortex showed increased activation for both or either type of cue for causation. Bilateral prefrontal cortical activation was seen with tasks requiring logical inference to make a judgement of causation. These results add to understanding of how the visual system extracts causal information from spatial and temporal cues and further suggests that causality arising from inference and real world knowledge, rather than from direct perceptual experience, may correspond to bilateral activation of the prefrontal cortex.
Fugelsang et al. (2005) state that their findings confirm theories that extraction of causality is inherent in the visual system, akin to the innate Gestalt principles of completion of partial contours and perceptual grouping based on similarity or spatial proximity. In addition, "perception of causality appears very early in human life and is culturally invariant" supporting the view that the "visual system may be specially tuned to recover causal structure from the environment" (Fugelsang, et al., 2005, p. 41). Several other studies also support the hypothesis that extraction of causal structure is an innate property of the brain structures of the visual system (Blakemore, et al., 2001; Fonlupt, 2003; Roser et al., 2005). Perception of causality is immediate like perception of motion or perception of faces suggesting innate mechanisms for perception of causation. Some authors have suggested that the detection of causality may even be served by a specialized brain module for recognizing and understanding causality (Leslie & Keeble, 1987; Scholl & Nakayama, 2002). Furthermore, infants are sensitive to physical causality at only 6 months of age (Leslie & Keeble, 1987; Oakes, 1994).
The exact brain areas activated during detection of causation depends on a number of factors. Judgments of causality that require integration of a working hypothesis with relevant data activate neural tissue in prefrontal and occipital cortices. In addition, evaluation of data that is consistent with a plausible causal theory, was reported to recruit neural tissue in the parahippocampal gyrus. By contrast, evaluation of data inconsistent with a plausible causal theory recruited neural tissue in anterior cingulate, left dorsolateral prefrontal cortex, and precuneus, suggesting a neural mechanism by which working hypotheses and evidence are integrated in the brain (Fugelsang & Dunbar, 2005). These empirical studies support the theory that, over the course of evolution, information about cause-effect relations has been genetically internalized into the brain. Innate understanding of causality is a highly adaptive component of intelligence in humans and animals. For example, without an understanding of causality, the planning and execution of goal-directed action on the environment would not be possible.
Similarity
It is a fundamental property of the world that things show similarity to other things in various features and at varying levels of abstraction (i.e. "There is nothing new under the sun."). An apple is similar to other apples and apples are similar to many other fruits. A wolf and a panther are both dangerous predators. All birds have beaks and wings. Prey animals frequent water holes. Both an obsidian knife and an obsidian arrowhead can penetrate a prey animal's flesh. Fast flowing rivers are dangerous to cross. Similarities among things in the world hold important adaptive information. Similarity allows categorization, prediction, and inferences based on generalization. If in the past I have witnessed fast flowing rivers wash people away, then when I encounter another fast flowing river, I can use the similarity to make the prediction that crossing this river could be dangerous, so I adjust my behavior accordingly and perhaps save my life.
This seems so obvious that we can't imagine not behaving so. It would be very stupid, not to be able to take advantage of similarities between past experiences and a current situation, especially when it might mean life or death. And that is exactly the point. It would be stupid, but we and other animals are not stupid, but the reverse, we are intelligent. But where did this ability for generalization come from? It is in our brain so firmly that we readily make such judgments of similarity, and generalize from them to make predictions, and we do this with such ease that we don't even think about it. But in neuroscience, it is those things that we do with greatest ease that most require explanation.
Extraction and exploitation of information such as this, contained in similarities, is an extremely powerful adaptive opportunity which natural selection could not have missed during the course of brain evolution. The fact that generalization and even categorization are found in a wide range of animal species (Soto & Wasserman, 2015) shows that these species are capable of using similarities in the environment to improve adaptation and biological fitness. This suggests that the adaptive information present in similarities drove the evolution of brain mechanisms, from early on, that could efficiently extract this information and put it to adaptive use, at least in birds and mammals, and perhaps in other animal groups as well (e.g. generalization has been found even in some insect species; see below).
Thus, “evolutionary/genetic internalization” (Shepard, 1994, p. 26) by natural selection of similarity, as an abstract, ubiquitous property of the world, provides another innate principle of how intelligence and cognition are organized. Just like we and many other animals have a "cognitive instinct" to understand and to experience the world in terms of cause-effect, we have a cognitive instinct to detect and readily utilize similarities to organize adaptive behavior and improve biological fitness.
Both causality and similarity are properties of how the physical world is put together and both have been incorporated into evolved mechanisms of intelligence in the brain. This makes sense. The brain must organize movement, behavior, in the physical world on our planet and therefore brain mechanisms must be effective in guiding behavior in that physical world. To do so, intelligence must have evolved to reflect and be organized around the general principles of how this physical world works--it operates by these universal principles, cause-effect and the fact that things in the world are naturally similar to other things in ways that provide organisms with information that is important for adaptation.
Genetic internalization of similarity as a general feature of the world created a disposition in human and animals brains to “expect” and to find similarities in the world, to group things by similarity into categories, to readily match new instances to the appropriate category based on similarity, and to infer properties of new instances of a category based on knowledge of properties of the category as a whole. The cognitive processes of generalization, concept formation, categorization, inductive reasoning, categorically based inference, and categorical logic, all emerged as a family of related functional properties of the brain as natural selection fine-tuned the genetic/evolutionary internalization of similarity as a fundamental relational regularity in the world (on this basis, it is expected that these cognitive abilities should be correlated). These observations are reflected in “what William James (1890/1950) called ‘the very keel and backbone of our thinking’: sameness. The ability to evaluate . . . similarity . . . is clearly the sine qua non of biological cognition, subserving nearly every cognitive process from stimulus generalization and Pavlovian conditioning to object recognition, categorization, and inductive reasoning” (Penn et al., 2008, p. 111). Note that many of these psychological capabilities are the same ones identified earlier in the definition of general intelligence from Gottfredson (1997).
As noted above, similarities hold important adaptive information. Similarities bring order out of chaos. They allow inference and prediction. If you know about one leopard as a member of a category of dangerous things, then you know about many leopards, perhaps all leopards, even those you have yet to encounter. If and when you encounter a leopard again, you already know much about it that can be put to use to organize your response, your behavior, and that knowledge that you gain from an inference about this new leopard, based on the category to which it belongs, may save your life. Evolution has "tuned" our brains to find similarities among things in the world--along with genetic internalization of causation, another exceedingly powerful property of general intelligence as described earlier. As noted above, the inborn disposition of the brain to find and record similarities in the environment guides us (our brains) to form categories and concepts of varying degrees of abstraction which embed adaptive information within them (e.g. all saber-toothed cats are dangerous; pieces of flint are sharp making them a good source of cutting and scraping tools; all Datura flowers are poisonous; things with feathers that fly are all birds; situations where someone is treated unfairly are all examples of injustice; all atomic particles with a negative charge are electrons, and so on). Categories and concepts are one way in which the brain captures the information contained in similarities in the world and forms knowledge structures that reflect and exploit one form of order in the environment, that order that arises from similarity, thereby improving the guidance of movement, increasing successful adaptation by intelligent action.
Recall that Shepard argued that “the evolutionary internalization of universal regularities” (1994, p. 26) included “three-dimensional, locally Euclidian space,” one-dimensional time with a "thermodynamically conferred unique forward direction," and cycles of light and dark. The genetic internalization of these regular features of the world underlies important components of our ability to understand the world. However, in addition, Shepard has also argued that there has been "evolutionary internalization" of the recurrent fact of the world that "objects having an important consequence are of a particular natural kind . . . however much those objects may vary in their sensible properties . . . " (Shepard, 1992, p. 500). In other words, things can be categorized on the basis of their consequences, regardless of their superficial physical properties. This more abstract form of similarity leads to grouping things which otherwise differ in surface ways but which share the same category of effects, for example, all large predators including bears, wolves, sharks, etc. are dangerous. Other things with very different superficial features may all be good to eat, or poisonous, or have other important adaptive consequences by which they can be grouped. This kind of more abstract categorization, based on similarities in effects, allows a higher level of behavioral guidance by the brain. Some of these categories and the behavioral responses to them may even be innate. For example, feces or spider-like things may innately cause feelings of disgust or fear and lead to withdrawal and avoidance behavior, thereby protecting the organism from potential harm from pathogens or a poisonous bite. The claim that "objects having an important consequence are of a particular natural kind" is basis for Shepard's “universal law of generalization” (Shepard, 1987b), a law which has been verified in diverse species including insects (e.g., see Cheng, 2000).
A quick review is important here. The psychological process of generalization exemplifies a basic property of human and animal minds--that they tend to form groupings, categories, based on similar instances (Broschard, et al., 2021), and then, exploiting these groupings, the mind/brain uses generalization to make inferences or projections about new instances of a category. For example, if you have encountered a number of cats in your past, you will form a grouping in your mind, a category, "cat," and when you see another creature in the future similar to previous cats you have encountered, you can already make a number of inferences about this new instance of the category--e.g. it has claws, it purrs, it eats mice, etc. You can infer these properties even though you have just encountered this particular cat only a few moments before. Because of categorization and generalization, you can make inferences and predictions about this new creature which you have recognized as a member of the category, "cat," based on similarities among all cats. This is an exceedingly important component of our thinking processes and our intelligence. We use this type of process much of the time when we are engaged in "thinking" and "reasoning."
Recognizing similarities among things, and using those similarities to make useful inferences and predictions has exceedingly beneficial effects on survival and sometimes directly upon reproduction. For example, chimpanzee males of lower status have been observed to steal a mating opportunity usually not available to them, when a more dominant male is sufficiently distracted. When a lower status male observes more dominant males fight these males are usually distracted from other matters. If in most situations where dominant males are fighting they are distracted from their watch over their favorite females in the group, and if a lower status male chimp recognizes a similarity between a current fight between dominant males and past such battles, and if, based on the similarity, he infers that during a current fight between dominant males, he likely has a mating opportunity (especially if he is quick), he may successfully mate with a desirable female that he would not otherwise have access to. Males who can make inferences like these based on similarity have a higher reproductive rate than males who cannot make such inferences.
Ancestral humans who recognized similarities between a new animal, never encountered before, and predators encountered in the past, can put this new animal in the category "dangerous predator" and then infer that this new animal is probably also dangerous and should be avoided. The adaptive value of this inference based on similarity is immediate. Imagine an ancestral human who had brain organization incapable of recognizing similarity at all and consequently could not form categories and concepts, and therefore could not make any inferences based on similarity. In this hypothetical case, where similarity to previous predators holds important survival information, clearly survival is at stake, and if one had a brain incapable of exploiting the survival information in similarity then one's chances of seeing another day are vastly reduced and the genes responsible will probably soon be eliminated (an instance of natural selection at work).
Research suggests that category formation depends on the prefrontal cortex in humans, and on the pre-limbic cortex in rats (Broschard, et al., 2021). Studies of brain damaged humans also implicate a subcortical involvement of the striatum. Specifically, Lagarde, et al. (2015) report involvement of "a prefrontal-striatal loop in abstract categorization, and more particularly, the involvement of the head of the caudate nuclei and left frontal lobe in access to abstract representations in verbal concept formation." To emphasize the point once more, the ability to form groupings, or categories, based on similarity, and to then make inferences about newly encountered instances of the category, is a powerfully adaptive property of the brain's operations. Categories and concepts based on similarity permit our brains to make inferences about newly encountered instances of the category or concept, like those in the examples above. Categorization, formation of concepts, and inferences based on similarity are key features of general intelligence. All of these cognitive abilities can be understood as arising from the genetic internalization of similarity relations into brain organization by natural selection (Koenigshofer, 2017).
Predictive Relations (Predictive Co-occurrence of Events)
In addition to causality and similarity, it is also an enduring property of the world that some things regularly predict the occurrence of other things (a growling bear predicts an impending attack, sudden heavy rainfall predicts possible flash flooding, an approaching range fire predicts potential danger, a water hole on the plains of Africa predicts that prey animals will be near, and so on). Clearly, predictive relations are present everywhere in the environment. It is also clear that predictive relations among things in the world hold exceedingly important adaptive information. This source of adaptive information is far too important for natural selection to have missed it. Thus, we should expect that natural selection must have organized brain systems that capture, not only causality and similarity, but also the predictive relations between events in the world. Using Shepard's (1992) terminology, we should expect that over the course of brain evolution, there occurred the "genetic internalization" (Shepard, 1992) of mechanisms that capture predictive relations between events. These mechanisms underlie another central component of general intelligence in humans and many animals.
The brain evolved for prediction (Clark, 2013, 2016; Nave, et al., 2020). The brain can be characterized as "a probabilistic prediction engine" (Nave, et al., 2020, p. e1542). It is not only evolved for prediction, but to implicitly make probability assessments about how likely its predictions about future are to be true. Natural selection for brain organization that captures the adaptive information in predictive relations has predisposed us (i.e. tuned our brains, and the brains of other animals) to find and to exploit the adaptive information held in predictive relations, an exceedingly adaptive property of general intelligence. Predictive relations exist whenever specific events, objects, or situations consistently co-vary or correlate--whenever one thing consistently follows another; for example, think of the CS and US in classical conditioning. After conditioning has occurred, the CS predicts the US. This implicit prediction then controls behavior--the dog salivates immediately after the CS is presented in anticipation of coming food, a response that prepares the dog for the expected arrival of the US (e.g. the dog drools, and this saliva prepares the dog to swallow and digest efficiently if the expected food follows). As discussed in the chapter on learning and memory classical conditioning specifically involves the cerebellum, whereas associations, in general, between co-occurring events appears to depend on long-term potentiation (LTP), and corresponding changes at synapses in hippocampus (Oishi, et al., 2019), cerebral cortex (Daw, et al., 2009; De Pasquale, et al., 2014) and other brain regions.
The adaptive significance of this property of general intelligence is hard to overestimate. An innate predisposition built into the brain to search for predictive relations in the environment and to exploit the adaptive information in those relations is an exceedingly powerful cognitive tool for understanding the world. Using this information to make biologically important predictions (a low growl in the nearby brush may be soon followed by an attack from a large predator), to organize behavior serves biological fitness (survival and reproduction of one's genes). To understand the powerful effect on adaptation of an innate disposition to recognize and exploit predictive relations between events in the world, just imagine the crippling effects on survival if this ability were absent in animals and humans. If you had no knowledge of what follows what in the environment, the world would seem a chaotic jumble of unrelated events. In such a world, it would be very difficult if not impossible to organize adaptive action to survive the demands of a complex environment. One might speculate, and this is only speculation, that something like this might be how some persons with autism experience the world; perhaps the inability to understand cause-effect or to recognize similarities in order to form categories and make sensible inferences, or inability to recognize predictive relations in the world, or perhaps loss of all three of these abilities might be responsible for some of the symptomology of people who are very severely autistic. The world would seem to lack any order at all without these cognitive abilities. Ordering movement into adaptive patterns would be exceedingly difficult if not impossible.


Figure \(\PageIndex{8}\): Contrasting behaviors to satisfy water needs in two species. The woman uses invention and technology, two products of human intelligence, to provide a more consistent and higher quality water source. (Images from Wikimedia Commons; Elephants drinking; https://commons.wikimedia.org/wiki/F...k,_Kenya_4.jpg ; by CT Cooper; licensed under the Creative Commons Attribution 3.0 Unported license. Woman pumping well; https://commons.wikimedia.org/wiki/F...an_Working.jpg; by Osaba Gerald; licensed under the Creative Commons Attribution-Share Alike 4.0 International license; caption by Kenneth A. Koenigshofer, PhD).
Intelligence: Cognitive Instincts and "the Infinite Use of Finite Means"
Recall this quote from above: "Evolutionary psychologists sometimes argue that a class of situations must be relatively narrow to exert consistent selection pressure, but this claim is insufficiently justified. Any regularity in the environment can exert selection pressure if it poses a challenge or an opportunity to the organism . . . (Kaufman, et al., 2011, p. 213)
In the case of the abstract relational regularities which drove the evolution of general intelligence, the adaptive opportunity is difficult to overstate. The genetic internalization by natural selection of fundamental information about these abstract relational regularities in the environment has equipped the mind with an exceedingly powerful set of cognitive structures--a set of inborn, general, rule-like principles of how the world works. Deployment of this inborn, implicit “understanding” about causality, similarity, and predictive relations gives general intelligence (g) its adaptive punch because it permits innovative and improvisational solutions to an enormous variety of adaptive problems. This is an example of what Steven Pinker (2007) calls the “infinite use of finite means,” applying a finite set of rules to understand and to solve an enormous variety of adaptive problems and to exploit the diverse opportunities in the environment. We see the power of human general intelligence in the stunning progress in human cultural and technological development from prehistoric times to the present. The existence of general intelligence (sometimes called "fluid intelligence") contradicts the traditional view sometimes ascribed to evolutionary psychologists that there are specialized modules in the brain for every problem type that ancestral humans had to solve (this is the theoretical assumption of the "massive modularity" of the human mind which leaves no room for more general cognitive processes including general intelligence--see Lloyd & Feldman, 2002; but for an alternative view held by most evolutionary psychologists, "soft modularity," which does allow for some general cognitive processes, see Ellis & Ketelaar, 2002). However, arguably there is no real contradiction if one understands that general intelligence is actually composed of several adaptive specializations that have genetically internalized the abstract properties of causality, similarity, and predictive relations. It is the internalization of these relations, because they are universally present in the world, that gives the mistaken impression of "general process" mechanisms which in fact are actually evolved adaptive specializations to these three specific abstract relations found in the structure of the world (Koenigshofer, 2017).
Genetically internalized information about causality, similarity, and predictive relations as general features of the structure of the world creates a scaffolding of instinctual knowledge which guides the learning of specific details of these relations in one's own particular environment, an environment full of specific, perhaps even unique, instances of these pervasive relational regularities of the world (Koenigshofer, 2017). For example, understanding causality as a general principle of the world allowed our ancient ancestors to learn that fire could be used to cook meat making it easier to digest; that setting well-placed fires on grasslands could be used to heard prey animals toward an ambush to facilitate the hunt; that some stones could be shaped into spear heads and tips of arrows; and that skins of animals could be taken and used for clothing. This same instinctual knowledge about causality permitted modern humans to learn how to make and use concrete to lay building foundations; to learn that fertilizers increase crop yields; that rising interest rates cause stock prices to fall; and that sunlight could be converted to useable electricity.
Innate knowledge about cause-effect as a general property of the world allows humans to analyze complex causal relations to solve a wide range of adaptive problems with creativity and innovation unprecedented in the animal kingdom. For example, while most animals seek shelter in vegetation, or in caves, or other features of the natural habitat, humans build castles, houses and skyscrapers. While other animals walk and run or fly to move from one place to another, humans invented the wheel, engines, and steel and now ride in cars, trains, planes, or even a Blue Origin space capsule. While most animals hunt for water and food, humans have invented plumbing, water companies, agriculture and supermarkets. While animals fight with teeth and claws, humans have invented and used spears, guns, and nuclear weapons. These intellectual achievements involve a general understanding of causality, similarity, and predictive relations as well sophisticated understanding of three dimensional spatial relationships, ability for visualization, and culturally transmitted knowledge accumulated and transmitted over generations, primarily and most efficiently by language.
This view that we are born with genes which give our brains innate knowledge about many things in the world is a radical departure from the view that the mind is a blank slate at birth. This conception of the mind was promoted by a group of influential 18th century philosophers known as the British Empiricists (such as John Locke). This view of the human mind has been carried forward all the way into 21st century social science and philosophy. Evolutionary psychologists, Cosmides and Tooby (2007), call this old empiricist view of the mind the Standard Social Science Model (SSSM). We now know that this blank slate view of the mind/brain (SSSM) is wrong. For example, as mentioned earlier, Alison Gopnik and her colleagues at Berkeley (Gopnik and Sobel, 2000; Gopnik, et al., 2001; Gopnik, et al., 2004) have found that children, at a very early age, readily discover cause-effect and predictive relations between events in the laboratory, supporting the view that the human brain is genetically predisposed to understand abstract relations such as causality and predictive relations among things in the world (Koenigshofer, 2017). Evolutionary psychologists, Cosmides and Tooby, call the blank slate view of the human mind (the SSSM) "biologically naive," "radically defective," and an outdated, theoretical leftover from the days of radical behaviorism.
Cognitive Instincts and Intelligence
As was described in the theoretical account above, the brain has genetically internalized by natural selection crucial information about regularities in the world of varying degrees of concreteness and abstraction. This equips us and other animals with inborn, instinctual knowledge about many enduring facts about the environment. Snakes, spiders, and heights have been consistently dangerous to humans generation after generation so natural selection has built this across-generation fact into our brain circuitry in the form of fear circuits readily activated by these stimuli. Certain facial and body forms in the opposite sex are consistently associated over generations with health and high reproductive potential so we have brain circuitry that innately attracts us to these body and facial forms. Three-dimensional space, the forward progression of time, cycles of light and dark, and Shepard's universal law of generalization have been "genetically internalized" into the brain over eons of evolution. And, as we have just discussed, natural selection has "genetically internalized" information about some quite abstract and universal regularities of the world: causality, similarity, and predictive relations. As a consequence, we are innately predisposed to understand cause-effect, the three-dimensionality of space, the progression of time, similarity relations used to form categories and concepts, to fear snakes and spiders, to seek particular traits in sexual partners, to look for and find predictive relations among events in the world, to understand the minds of others, to protect our young, to form and maintain friendships, and so on. This means that we are born with many genetically encoded "cognitive instincts" which make up much of our genetically inherited intelligence as a species, the consequence of eons of natural selection.
Details of how natural selection could bring about the genetic internalization of abstract relational regularities of the world (causality, similarity, and predictive relations) are described elsewhere (see Koenigshofer, 2017). Suffice it to say, that adaptive problems and opportunities, though variable or even “novel” in details, are nevertheless invariant in common abstract, relational structure. That recurrent structure provides stable selection criteria over generations for genetic internalization. Since natural selection can only operate over many generations, the specific details of individual cases of each of the abstract relations (cause-effect, similarity, predictive relations) drop out over generations. This leaves only enduring, across-generation, "distilled" abstract relational regularities (independent of specific concrete contents) to act as selection criteria for the genetic internalization of these relational regularities of the world into brain operations. This process has apparently taken place not only in humans, but also in some number of non-human animals as well, since general intelligence has also been identified in a variety of non-human animal species (Bird & Emery, 2009a, 2009b; Reader, et al., 2011), as well as in humans.
What makes humans so smart?
According to the theoretical views explained above, genetic internalization of these enduring, across-generation, abstract, relational regularities of how the world works drove the evolution of flexible general intelligence in humans and many other animal species. But what makes humans so smart compared to other species on the planet? One key difference between human intelligence and intelligence in non-human animal species is the much higher degrees of abstraction that humans are capable of representing, compared to other animals. The ability of the human brain for high levels of abstraction, setting it apart from the brains of other species, is likely due to the greater complexity of circuits in human cerebral cortex compared to other mammals (recall that only mammals have six layered cerebral cortex and approximately 150,000 cortical columns--discussed above--while non-human mammals have significantly fewer). Humans easily detect highly abstract similarities in the environment compared to non-human animals and are capable of forming highly abstract categories and concepts based on these abstract similarities in properties or functions (Koenigshofer, 2017; Penn et al., 2008). For example, Penn et al. (2008) found that the use of causal principles and similarity assessments of high degrees of abstraction is a distinguishing feature of human cognition and problem solving. As they state: “Even preschool-age children understand that the relation between a bird and its nest is similar to the relation between a dog and its doghouse despite the fact that there is little “surface” or “object” similarity between the relations’ constituents” (Penn et al., 2008, p. 111).
Research in molecular genetics suggests one possible explanation for how human ability for high levels of abstraction may have come about. Pollard (2009), comparing human and chimpanzee genomes, found “massive mutations” in humans in the “DNA switches” controlling size and complexity of cerebral cortex, extending the period of prenatal cell division in human cerebral cortex by several days compared to our closest primate relatives. Research using artificial neural networks suggests that increasing cortical complexity leads to sudden leaps in ability for abstraction and rule-like understanding of general principles (Clark, 1993), lending further support to the hypothesis that superior ability for abstraction due to cortical complexity may be the key component explaining differences in general intelligence between humans and nonhuman animals (see module on artificial neural networks in this chapter).
The frontal cortex is involved in concrete rule learning. More anterior regions along the rostro-caudal axis of frontal cortex support rule learning at higher levels of abstraction. These abilities for high levels of abstraction may involve anterior dorsolateral prefrontal cortex in humans (Kroger et al., 2002; Reber, Stark, and Squire, 1998). Along with human language, the exceptional capacity of the human brain for abstraction may explain the unusual achievements of the human species ranging from agriculture, technology and science, to the invention of complex economies and governments (Koenigshofer, 2016, 2017).
Furthermore, cultural transmission of learned information in humans, ranging from books and educational institutions to film and the internet, allows each generation to profit from the accumulated knowledge of prior generations (it might even be said that cultural transmission is a key specialty of the human species). Cultural transmission, along with cooperative problem solving and development of experts in different fields, creates a kind of group intelligence not possible in other species. Of course, human language, spoken and written, plays an enormous role in these shared cognitive processes among humans, perhaps explaining at least in part why language evolved. These considerations bring to the forefront the role of culture in human genetic evolution. Gene-culture coevolution theory proposes a multidirectional coevolution between genes and culture in which culture and human created artifacts such as tools, weapons, clothing, and pottery have driven the last 100,000 years of human evolution (Lloyd & Feldman, 2002; Durham, 1991). This theory also assumes an important role for language in cultural transmission of learned knowledge and behavior. It also postulates existence of hominin biological changes that facilitated development of sophisticated language in humans (Lloyd & Feldman, 2002).
.jpg?revision=1&size=bestfit&width=612&height=390)
Figure \(\PageIndex{9}\): Scientists seek cause-effect, similarity, and predictive relations between variables. Scientific methods are an example of human general intelligence at its best. Alison Gopnik (2010, 2012) at UC Berkeley has found that young children engage in logical analysis of causation, teasing out causes from non-causal factors, in much the same way that scientists do. (Image from Wikipedia, https://commons.wikimedia.org/wiki/F..._reaction).jpg; by Alenakopytova; licensed under the Creative Commons Attribution-Share Alike 4.0 International license).
Another factor that may contribute to the superior intellectual capacities of humans is the sophisticated control systems in the human prefrontal cortex. Miller and Cohen (2001) proposed that “cognitive control stems from the active maintenance of patterns of activity in the prefrontal cortex that represents goals and means to achieve them. They provide bias signals to other brain structures whose net effect is to guide the flow of activity along neural pathways that establish the proper mappings between inputs, internal states, and outputs needed to perform a given task.” On their view, the prefrontal cortex (PFC) guides the flow of neural activity in relevant brain areas, which allows for cognitive control of behavior. As they state: "depending on their target of influence, representations in the PFC can function variously as attentional templates, rules, or goals by providing top-down bias signals to other parts of the brain that guide the flow of activity along the pathways needed to perform a task" (Miller & Cohen, 2001).
Goldman-Rakic (1996) proposed that the prefrontal cortex represents information not currently in the environment, creating a "mental sketch pad," to hold visual images in working memory, to represent plans, and to focus attention to intelligently guide thought, action, and emotion. This includes the inhibition of distracting thoughts, actions, and feelings. Although such control does exist to some degree in some non-human animals it is less developed and less influential over behavior than in our own species. The prefrontal cortex is highly interconnected with much of the brain, including extensive connections with other cortical, subcortical and brain stem sites. The dorsal prefrontal cortex is especially interconnected with brain regions involved with attention, cognition and action, while the ventral prefrontal cortex interconnects with brain regions involved with emotion. According to Striedter (2005), the prefrontal cortex (PFC) of humans consists of two functionally, morphologically, and evolutionarily different regions: the ventromedial PFC (vmPFC) composed of the ventral prefrontal cortex and the medial prefrontal cortex present in all mammals, and the lateral prefrontal cortex (LPFC), composed of the dorsolateral prefrontal cortex and the ventrolateral prefrontal cortex, present only in primates.
Mental Models of the World: Capturing Environmental Regularities for Prediction and Planning for Future
Think for a moment about the complexity of the environment that animals like us must adapt to. We live in a world that consists of inanimate and animate objects, including our own body, that move and change in space and time. Forces that operate on objects can be physical like gravity and the laws of motion, while for people and animals, additional forces acting on them can be social, emotional, or generated by physiological or other needs and motives. The relations among things can be spatial (e.g. the lion is approaching me from my right), temporal (e.g. right after the bell rang a pellet of food appeared), causal/logical (germs cause disease), predictive (one event signals that another event is about to follow), social (interactions among conspecifics), and can reflect similarity (a bear and a wolf are both large predators) and differences (wolves hunt in packs, bears don't), as well as other entities of even greater levels of abstraction.
To successfully navigate this incredibly complex world and its causal and other contingencies, the brain must have accurate neural representations or mental models, inclusive of all of these objects, forces, and relations, in order to guide adaptive behavior. Consequently, evolution has equipped us with a brain that generates neural representations (mental models or "cognitive maps") of things (including ourselves, other people, animals, processes, etc.), the relations among them, and the forces that operate on them in three-dimensional space and time (see Behrens, et al., 2018). Many psychologists and neuroscientists have recognized the importance of mental models in the intelligent organization of behavior. "Mental models are psychological representations that have the same relational structure as what they represent. They have been invoked to explain many important aspects of human reasoning, including deduction, induction, problem solving, [and] language understanding, . . . [and they are] also valuable for providing new understanding of how minds perform abduction, a kind of inference that generates and/or evaluates explanatory hypotheses" which focus on explaining causal mechanisms (Thagard, 2010, p. 447). "[T]hese models underlie deductive, inductive, and abductive reasoning yielding explanations" that permit us to understand the world (Khemlani, et al., 2014) and then to use this understanding to predict, to plan, and to manipulate objects and events in the environment to human advantage. In an important sense, the use of mental models to generate adaptive behavior encompasses much of what we mean by intelligence.
Neural Coding and Mental Models
Mental models are thought to be generated by the activities of large populations of neurons in the brain. One theory of how this is done proposes that complex ideas, concepts, perceptions, memories and other mental structures are represented by specific patterns of neural firing, over different regions of brain, in enormous numbers of neurons, each "tuned" to fire to a specific feature or to a specific value of a feature or property of the world (recall Hebb's idea of "cell assemblies" discussed in the chapter on learning and memory). Neuroscientists know that this sort of coding of properties exists in many parts of the brain. For example, recall from the chapters on vision and other sensory systems that individual neurons in the visual cortex are "tuned" (i.e. have receptive fields) to specific visual properties such as the orientation of linear borders and their direction of movement, while some neurons in somatosensory cortex, for example, respond to specific angles of specific joints of the skeleton to code limb positions, while other respond to tactile stimulation of specific areas (dermatomes) of skin, while still other neurons in auditory cortex fire to a specific narrow range of frequencies. Paul Churchland, at the University of California, San Diego, postulates that complex stimuli such as human faces or even highly abstract concepts and ideas such as scientific theories may be neurally represented in the brain by complex patterns of firing in enormous populations of neurons, each tuned to fire neuron potentials to specific features and magnitudes of features found in the world or generated by creative recombination of features in imagination.
This kind of coding can be conceptualized as long strings of numbers (each number is referred to as a "scalar," and the entire string of scalars is called a "coding vector") in which each number (scalar) in the string (the vector) corresponds to the firing rate of a specific neuron in a large population of neurons involved in the coding of a perception, a concept, an idea, or a complex set of ideas such as a scientific theory. This means that coding vectors for a single idea might be exceedingly long, containing thousands or millions of scalars in a single coding vector, each such scalar representing the firing rate of one of thousands or even millions of neurons involved in the coding of a single concept, idea, or complex of interrelated ideas (see Churchland, 1985, 2013).
Consistent with this view, the coding capacity of a single human brain with its 100 to 200 billion neurons is staggering, well within the parameters suggested by Churchland. As Thagard (2010, p. 450) explains: "A population of neurons represents something by its pattern of firing. The brain is capable of a vast number of patterns: assuming that each neuron can fire 100 times per second, then the number of firing patterns of that duration is (2 (100))100000000000, a number far larger than the number of elementary particles in the universe, which is only about 1080. A pattern of activation in the brain constitutes a representation of something when there is a stable causal correlation between the firing of neurons in a population and the thing that is represented, such as an object or group of objects in the world."
According to mental model theory, the enormous coding capacity of the human brain permits the human mind to represent concepts, ideas, theories, and other components of human knowledge of great subtlety and complexity, including theories in science about hidden causes of things not directly observable (Churchland, 1985, 2013). "[M]ental models operating at various degrees of abstraction are invaluable for high-level reasoning. . . you can build simplified but immensely useful models of past and future events, as well as of events that your senses do not enable you to observe. Hence science uses abductive inference and conceptual combination to generate representations of theoretical (i.e. non-observable) entities such as electrons, viruses, genes, and mental representations" (Thagard, 2010, p. 457).
Using Churchland's concept of vector coding, each neural representation of a perception, a concept, an abstract relation, a thought, or other mental activity could be conceptualized as a specific, very long, coding vector, perhaps consisting of thousands or millions of scalars corresponding to the firing rates in an equal number of neurons which are involved in the coding of that particular mental activity. By this reasoning, mental models of the world would involve patterns of firing over space (neural space; i.e. areas of brain) and time (temporal patterning) in enormous numbers of neurons, corresponding to coding vectors of vast length changing their scalar values and composition moment to moment. This is hard to imagine, but perhaps this is the scale on which neural coding of mental events, such as concepts, relations, ideas, and so on takes place--millions upon millions of neurons firing in ever changing spatial-temporal patterns of neural activity distributed across many regions of brain at any one time.
Brain Mechanisms, Mental Representations, Relational Reasoning and Intelligence
Research suggests that reasoning requires coding involving neurons in prefrontal cortex. "Reasoning depends on the ability to form and manipulate mental representations of relations between objects and events. . . [and] the integration of multiple relations between mental representations . . .[However,] patients with prefrontal damage exhibited a selective and catastrophic deficit in the integration of relations. . . . The integration of relations may be the fundamental common factor linking the diverse abilities that depend on prefrontal function, such as planning, problem solving, and fluid intelligence" (Waltz et al., 1999, p. 119; fluid intelligence is another term for general intelligence, and is separate from one's store of learned knowledge). These results suggest that although mental models may involve many different brain areas acting together, the prefrontal cortex appears to play a special role in high level abstract reasoning requiring the integration of multiple relations. This may explain why damage in prefrontal cortex "leads to selective decrements in performance on tasks involving hypothesis testing, categorization, planning, and problem solving, all of which involve relational reasoning" (Waltz et al., 1999, p. 119).
In addition, because the ability to anticipate and prepare for future events improves chances of survival, evolution has favored brains that can predict future events (Clark, 2013; Nave, et al., 2020; Koenigshofer, 2017) including the future consequences of our own actions. Thus, according to mental model theory, intelligence involves formation by the brain of mental/neural models or mappings of the physical and social worlds and how they work. This idea is similar to Tolman's concept of "cognitive maps," in which he envisioned mental mappings of the environment even in rats learning a maze (Tolman, 1948). This concept has even been extended to understand the coding of social relationships (Schafer & Schiller, 2020). On this view, intelligence involves enormously complex neural representations, models, or mappings of the environment and how it operates, including its causal relations, from which it is possible to make causal inferences and to generate causal explanations. There is evidence that mental models involved in causal inference may depend upon localized regions of cortex. According to Khemlani, et al. (2014) "mental models for causal inference are implemented within lateral prefrontal cortex." Furthermore, planning future actions has also been associated by researchers with lateral prefrontal and parietal cortex. "Lateral prefrontal cortex, the anterior extent of the inferior parietal lobule, dorsal anterior cingulate, and anterior insula comprise regions of an extended frontoparietal control system" (Spreng, et al., 2015).
Recall Shepard's idea that there has evolved "a mesh between the principles of the mind and the regularities of the world." As discussed above, a primary characteristic of intelligence is that it predisposes animals and humans to discover and exploit regularities in the environment of varying degrees of abstraction. Similarities between things in the world help organize knowledge by the formation of categories (see module that follows on categories, concepts, and schemas). Inference from categories to specific instances of a category is one way to predict future outcomes (if this happens, then that is likely to follow), including the prediction of the effects of one's own future actions (if I do this, then that is likely to follow). Both of these kinds of predictions are examples of what we mean by "reasoning." The latter is critical for planning and the achievement of long-term goals.
Prediction allows us to plan ahead, to problem solve, and to achieve long-term goals, functions which depend upon prefrontal cortex (PFC). For example, one function of the prefrontal cortex may be to gather facts, memories, and information from various brain regions and then coordinate and organize this divergent information into a workable whole in working memory in the PFC in order to solve a puzzle or find a solution to a problem ( D'Esposito & Postle, 2015). However, an alternative theory is that the PFC is involved in selective attention to information which is localized in posterior brain regions such as occipital and parietal cortex. According to this view, the memories for relevant information are stored in the same regions activated during the initial sensory processing prior to memory formation (i.e. memories are stored in the sensory cortex). Information may flow in both directions from posterior regions of cortex to the PFC and from PFC to posterior regions of cortex during a visual cognitive task such as categorizing visual stimuli (Lara & Wallis, 2015). Some studies of sensorimotor processing have shown similar bidirectional interactions within the frontoparietal network (Siegal et al., 2015), a brain network which has been identified, using brain functional MRI (fMRI) and Positron Emission Tomography (PET), as a primary neural substrate relevant to intelligence (Colom et al., 2010; Hearne, et al., 2016; Jung & Haier, 2007).
The connection of parietal and frontal cortex to intelligence has been formalized in the Parieto-Frontal Integration Theory (P-FIT) of intelligence (Jung & Haier, 2007). According to Jung and Haier (2007), the P-FIT model of intelligence proposes that the dorsolateral prefrontal cortex, the inferior and superior parietal lobule, the anterior cingulate, and areas within the temporal and occipital lobes are crucially involved in intelligence. White matter regions (consisting of tracts of myelinated axons) such as the arcuate fasciculus are also involved (more details about this theory follow in a later module). This theory attempts to relate these brain areas and their functioning to individual differences among people in intelligence, as measured by standard intelligence tests (see module that follows on the measurement of human intelligence).
Representations of Space and Time as Components of Intelligence
As already discussed, similarity relations, predictive relations (event covariation), and cause-effect relations (Koenigshofer, 2017) are not the only environmental features which have been genetically internalized into brain operations by natural selection. In addition, as discussed above, there are fundamental properties of time and three-dimensional space (see Shepard, above) and a large number of other relatively concrete regularities of the world which have also been incorporated into the mental/neural models or cognitive maps of the environment created within our brains. Thus, we, like other animals, have specialized regions of the brain dedicated to time perception and others devoted to creation of three-dimensional representations of the space around us. Recall from above, the "place cells" discovered in the hippocampus of the rat. These are "tuned" to fire action potentials when the rat is in a particular place in its environment; recall that other cells in the cerebellum, hippocampus, and entorhinal cortex appear to be involved in representation of the passage of time (Irvy, et al., 2002; Schafer & Schiller, 2020). Natural selection favored brain circuitry capable of exploiting these universal regularities or invariants in the structure of the world, along with more specific regularities of the physical and social environments acquired through learning, allowing us to generate quite sophisticated mental models of how the world operates. These brain systems form neural groundwork for thinking and intelligence in humans and animals.
"From Reflex to Reflection" in the Control of Action
The better our mental/neurological models of the world are, the more accurately we can anticipate future events and thus prepare for them. The better we are able to prepare for future events, whether only moments or years ahead, the more successful, the more adaptive, our behaviors are likely to be. These mental models or cognitive maps are part of the sophisticated guidance systems that shape behavior into adaptive patterns. This allows humans and animals to meet the challenges, and to maximize the opportunities, presented by the environment. The result? Increased biological fitness measured by the successful reproduction of genes (Dawkins, 1976).
In fact, we can see an important evolutionary trend: the brain's increasing ability to anticipate future, beginning with reflexes (e.g. sexual reflexes anticipate coitus) and then classical conditioning, found even in simple animals, in which the organism learns to anticipate or expect the UCS (such as meat in Pavlov's experiments) a few seconds after the CS (the bell). Natural selection for prediction and the anticipation of future ultimately led to evolution of the ability for advanced mental planning and prediction in human thinking (Clark, 2013; Koenigshofer, 2017), sometimes involving anticipation of events decades or even centuries into the future (e.g. you may anticipate a well paying, interesting career years from now after you have completed college, and that mental image keeps you focused day to day on your goal and the intermediate steps to achieve it; scientists predicted global warning decades ago; Leonardo da Vinci in the 1400's anticipated invention of the helicopter by hundreds of years; Jules Verne anticipated space travel long before the first human space flight; the founders of the U.S. Constitution formed principles of governance which anticipated political behavior of people hundreds of years in the future to this day). This general trend in evolution toward increasing capacity of brains to guide behavior by anticipation of the future can be summarized in the phrase "from reflex to reflection" (Koenigshofer, 2011, 2016).
Recent research has identified a number of brain structures involved in mental representations of future states of the world, specifically of one's own personal future, goals, and plans, essential for intelligent control of behavior. Prospection is the ability to mentally represent the future. To reach a future goal it is essential to make plans which organize a sequence of actions leading to achievement of the future goal. Behaviors controlled by mental anticipation of imagined futures can be called prosponses to distinguish them from responses to stimuli in the present or recent past (such as a CS in conditioning) (Koenigshofer, 2016). According to Spreng et al. (2015), autobiographical planning involves personal plans directed toward real-world goals. These researchers found that autobiographical planning involves "synchronized activity of medial temporal lobe memory structures as well as frontal executive regions, . . . specifically, of the default and frontoparietal control networks. [The default network consists of] the medial prefrontal cortex (PFC), medial parietal cortex, including posterior cingulate cortex (PCC) and retrosplenial cortex (RSC), the posterior inferior parietal lobule (IPL), medial temporal lobes (MTL) [including hippocampus, amygdala, and parahippocampal regions], and lateral temporal cortex. . . [This network is activated] by self-generated thought and active across multiple functional domains including memory, future-thinking, and social cognition" (Spreng et al., 2015).
Earlier research revealed that the default network may be "a common 'core' network that" . . . "underlies both remembering and imagining." Functional MRI (fMRI) studies "reveal striking overlap in the brain activity associated with remembering actual past experiences and imagining or simulating possible future experiences" (Schacter, et al., 2012, p. 677-678). Clinical observations are consistent with this hypothesis from fMRI studies. For example, amnesic patients have difficulty imagining the future. Another study of amnesic patients with hippocampal damage revealed impairments when these patients were asked to imagine novel experiences (Schacter, et al., 2012). The abilities for imagining (see section below) and future-thinking are very important components of human intelligence.
How the World Works and Intelligent Control of Movement
Considering the complexity and variety of the many objects, forces and relations interacting in the physical and social worlds, it is not surprising that intelligence involves the integration of many brain areas acting together. As mentioned earlier, representation of three-dimensional space, including the position of one's own limbs, is dependent upon regions within the parietal lobe and one's location in local space is coded by hippocampal "place cells." Visual representation of objects depends upon the visual cortex in the occipital lobe, while visual recognition requires the inferotemporal lobe (lower temporal lobe, also known as IT cortex).
Human social interactions require other brain areas. A relatively new specialty called social cognitive neuroscience studies the brain systems involved in the information processing involved in social interactions, including some subcortical structures, several of which are part of the limbic system. Other researchers study neurocognitive adaptations underlying intuitive (or "folk") psychology, which humans use for inferring social causality in other humans, helping us understand their motives and predict their likely future actions. Others study intuitive (or "folk") physics, for innately inferring physical causality among inanimate objects in one's environment helping us anticipate future events (Baron-Cohen, et al., 2001; Kubricht et al., 2017). As already mentioned, developmental psychologists are studying young children's ability to use statistical regularities in the environment to discover the cause-effect relations between physical events (see Gopnik et al., 2001; Gopnik, 2012; Gopnik & Wellman, 2012).
To form mental models of the world to guide intelligent behavior, all of this information and information from additional sources must be combined. Rabini, et al. (2021) provide evidence that the precuneus area (see Figure 14.1.6) of the parietal lobe, which underwent a significant enlargement in fairly recent human evolutionary history, is critically involved in the mental combination of different mental concepts, in part, to create new more complex concepts and ideas, and perhaps to help generate integrated mental models of the environment. Among other things, the precuneus region (what is often referred to as the superior lobule of the parietal lobe which equates with the outer extension of the anterior and posterior precuneus) of the parietal lobe may have set the stage for the evolution of complex language and thinking by its involvement in the linking of concepts and ideas into increasingly complex ones in sophisticated human thought and complex cognition (Rabini, et al., 2021).
Planning for future ultimately involves action in the world and testing out the accuracy of one's mental models against feedback from the world. As Geertz (2016, p. 185) states:
The process of comparing models with perceptual input from the wider world is known today as “prediction error monitoring” (Frith 2007: 132ff.; Frith & Frith 2010). Our experience of the world is in fact our experience of the brain’s simulation of the world. The brain constantly predicts what is going on in the physical world, the body and, especially, the social world. These simulations are tested against the input of neurological mappings, perceptual input and social mappings. When it detects errors in its predictions, the brain attempts to improve its predictions.
Nave et al. (2020) express similar ideas. But attempts to improve predictions based on feedback from the world is only half the story. Not only does the brain attempt to improve its predictions, but it also directs the modification of behavior, based on feedback from the environment, to make corrections in action needed to keep behavior on track toward the achievement of an imagined goal. Continuous feedback from the environment about the effects of one's own behavior guides behavior toward imagined goals, with needed corrections along the way. For example, if you imagine going to law school and your grade on a midterm exam in one of your pre-law courses is below average, then this feedback from the environment that does not match your mental model of what is required to get accepted into law school can cause you to modify your study habits. This type of monitoring of the match or mismatch between an imagined goal and the actual effects of one's actions with respect to the goal, an important feature of intelligent action, involves interaction between the prefrontal cortex and the hippocampus. As Numan (2015, p. 323) states:
Action plans are essential for successful goal-directed behavior, and are elaborated by the prefrontal cortex. When an action plan is initiated, the prefrontal cortex transmits an efference copy (or corollary discharge) to the hippocampus where it is stored as a working memory for the action plan (which includes the expected outcomes of the action plan). The hippocampus then serves as a response intention-response outcome working memory comparator. Hippocampal comparator function is enabled by the hippocampal theta rhythm allowing the hippocampus to compare expected action outcomes to actual action outcomes. If the expected and actual outcomes match, the hippocampus transmits a signal to prefrontal cortex which strengthens or consolidates the action plan. If a mismatch occurs, the hippocampus transmits an error signal to the prefrontal cortex which facilitates a reformulation of the action plan, fostering behavioral flexibility and memory updating.
This sort of monitoring of the effects of one's actions comparing those effects to the effects desired to reach a goal is an essential part of the guidance systems for movement in complex animals, especially in humans, and is a central feature guiding intelligent action.
Above, we referred to cognitive instincts. Another of these instincts is emotional/motivational--what we commonly refer to as curiosity. As argued above, we are innately predisposed to understand cause-effect and to seek causal explanations of the world, "curiosity." As Thagard (2010, p.458-9) states: "abductive inference . . . leads people to respond to surprising observations with a search for hypotheses that can explain them. . . Brains make mental models through complex patterns of neural firing and use them in many kinds of inference, from planning actions to the most creative kinds of abductive reasoning." Understanding causality is not much use if you are not equipped with brain mechanisms which drive the motivation to know why things happen in your environment; that drive to explain, is the drive to identify causes. Knowing the causes of things allows better prediction, which leads to understanding of how to manipulate the environment to adaptive advantage--intelligence put to action.
General Intelligence: Adaptive Specializations to Abstract Relational Regularities
As described above, some of the most biologically important features of the world, of the environment, are abstract and relational. Recall that Spearman believed that g was most closely related to the "drawing out" of "relations and correlates,” abstract regularities of the world important in inductive and deductive logic, grasping relationships, inferring rules, and recognizing differences and similarities. Also discussed above, several of these broad, abstract properties of the world were central to the evolution of general intelligence and these have been genetically internalized into brain organization over the course of evolution. As previously noted, these include: 1) Causation: things/events in the world are governed by natural causal laws, and thus, causal relations among its elements are everywhere. Causality in the world holds important adaptive information. Innate knowledge about causality, by its genetic internalization, gives the mind ability to discover and exploit the biologically important information in causal relations, thereby improving adaptation; 2) Similarity relations: things/events in the world have similarities to one another in multiple dimensions and at varying degrees of abstraction (e.g. "there is nothing new under the sun"). Similarities hold important adaptive information extracted by the grouping of things into categories and concepts of varying degrees of abstraction; 3) Predictive covariation of events: things/events in the world covary or co-occur with one another in predictable ways, offering opportunity for prediction, and serving as an important cue to causality (Gopnik, et al., 2001; Gopnik & Wellman, 2012), both highly significant for successful adaptation. Genetic internalization of knowledge about these abstract relational regularities (causality similarity, and predictive relations) set the groundwork for the evolution of categorization, concept formation, generalization, causal and categorical logic and inference, and analogical reasoning, all of which are potent components of cognition and general intelligence (Koenigshofer, 2017).
From this perspective, general intelligence (Spearman's "g;" see discussion above and section 18.11) evolved as a collection of adaptive specializations (specific psychological adaptations to a particular class of environmental feature, relation, or adaptive problem; see Gallistel, 1995) to these universal, abstract relational regularities (causality, similarity, and predictive relations) of the world which repeatedly appear, generation after generation, in virtually all situations, adaptive problems, and adaptive opportunities in the environment. The adaptive information in causality, similarity, and predictive relations among events was far too significant to be missed by natural selection. Specialized brain mechanisms evolved to extract and utilize the information in these abstract relational regularities--forming the foundations of general intelligence in us and many other animals. The high levels of general intelligence that evolved in our species gave us ability to discover and understand general principles of how the world works that could be applied to a wide range of environmental problems and adaptive opportunities. General intelligence, becoming highly developed in recent human evolution (probably within the past 100,000 years; see Mellars, 2005), has allowed our species, unlike any other, to create the scientific method, systems of government and law, machines to do our work, modern medicine, agriculture, cities and nations, and space travel. Highly developed general intelligence combined with our capacities for cooperation and cultural transmission, both facilitated by language, set us apart from all other animals.
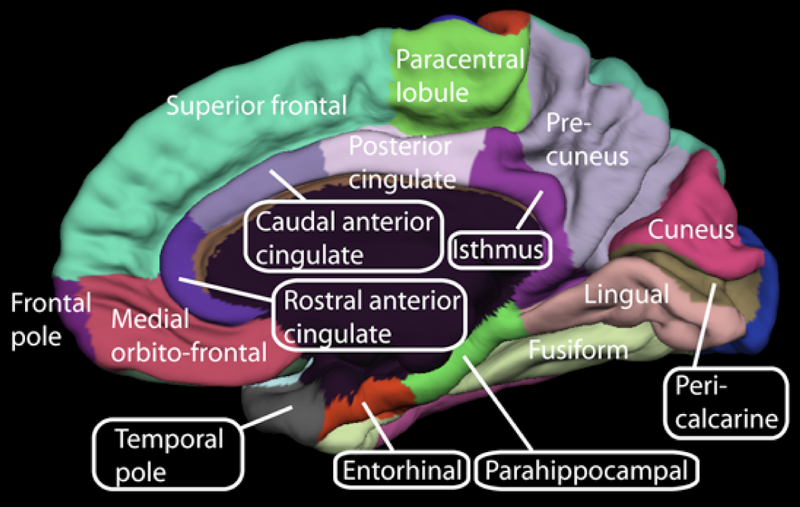
Figure \(\PageIndex{10}\): Artist's depiction of midline cross-section of human brain showing some of the structures involved in generating some components of intelligence (see text above); the hippocampus proper, buried in temporal lobe cortex, is not shown because it is located behind the plane of the midline view shown here. (Image from Wikimedia Commons; https://commons.wikimedia.org/wiki/F...tex_-_gyri.png; by Patric Hagmann et.al.; licensed under the Creative Commons Attribution 2.5 Generic license; caption by Kenneth A. Koenigshofer, PhD).
General Intelligence in Humans and Non-human Animals: Neural Correlates
Historically, psychologists have focused on ways of measuring intellectual abilities that are important for success in school. Traditional IQ tests were designed to measure these abilities. As discussed above, one important observation from research based on these intelligence tests is that measures of different intellectual skills are correlated: individuals that do well in one type of intellectual ability tend to do well in all types. These positive correlations among measures of different types of intellectual activity are known among psychologists as "the positive manifold." The "positive manifold" has been interpreted by many psychologists as strong statistical evidence (from the mathematical method, factor analysis; Spearman, 1904) for the general intelligence factor, "g," which exists in addition to more specific intelligences such as verbal fluency and visual-spatial abilities (see models of human intelligence in the next module).
Perhaps unexpectedly, recent research shows that general intelligence (ability to recognize "relations and correlates," according to Spearman,1904, 1925, who originated the concept) is found to some degree in many mammals and even in some birds (Emery & Clayton, 2004). Recent research shows that degrees of general intelligence among mammals is most strongly associated with "the number of cortical neurons, neuron packing density, interneuronal distance and axonal conduction velocity—factors that determine general information processing capacity (IPC), as reflected by general intelligence" (Dicke & Roth, 2016). These researchers compare IPC in various species. They report that: "The highest IPC is found in humans, followed by the great apes, Old World and New World monkeys. The IPC of cetaceans and elephants is much lower because of a thin cortex, low neuron packing density and low axonal conduction velocity. By contrast, corvid [rooks, crows, ravens] and psittacid birds [parrots] have very small and densely packed pallial neurons and relatively many neurons, which, despite very small brain volumes, might explain their high intelligence."
Figure \(\PageIndex{11}\): Size proportion of mature rodent and non-human primate brain as well as developing and mature human brains. Dorsal view of adult mouse (A), rhesus monkey (A), and human brain (B), as well as human fetal brain around mid-gestation (A) and at term (B). (A) The size of human fetal brain already at mid-gestation has reached the size of the adult rhesus monkey brain. Nevertheless, adult rhesus monkey brain is almost 100 times larger than brain of adult mouse. (B) The adult human brain is around 3–4 times larger than newborn brain which size reaches the size of adult chimpanzee brain. Note that the pattern of gyrification (cerebral gyri--the hill portions of the cortical folds) in human newborn brain is close to the one observed in adult. The inlet in the middle of the figure is the integrative photo of all brains shown in A and B that demonstrate their actual proportions (the right brain in the upper row of insertion is from fetus at the beginning of third trimester of gestation). Image and caption from Wikimedia. https://commons.wikimedia.org/wiki/F...00050-g004.jpg; by Ana Hladnik, Domagoj Džaja, Sanja Darmopil, Nataša Jovanov-Milošević and Zdravko Petanjek; licensed under the Creative Commons Attribution 3.0 Unported license).
An approach to individual differences among humans in intelligence involves efficiency of neural processing--higher IQ people show greater processing efficiency (Haier et al., 1992; Van Den Heuvel et al., 2009). Brain imaging studies show that higher IQ individuals show lower brain activation (as indicated by glucose metabolism) than lower IQ individuals when completing the same cognitive task, indicating that individuals with higher intelligence, as measured by IQ tests, do information processing more efficiently, with less expenditure of metabolic energy by the brain, compared to lower IQ individuals whose brains work harder (their brains expend more metabolic energy) at solving the same problem. Haier, et al. (1992, p. 415-416) conclude: "Intelligence is not a function of how hard the brain works but rather how efficiently it works."
Brain processing efficiency may be related to any number of factors: inherited brain circuitry configurations which are more "streamlined" as a result of more efficient neural pruning during brain development (Koenigshofer, 2011, 2016); better conduction over pathways connecting multiple regions of brain; "more long-distance connections that ensure a high level of global communication efficiency within the overall network" (Van Den Heuvel, et al., 2009, p. 7619); less engagement of brain areas not required to solve a particular problem or complete a particular cognitive task (Haier, et al., 1992); and/or as Hearne, et al. (2016) conclude, "higher global network efficiency is related to higher general intelligence measures" with "a key role for connections between prefrontal and frontal cortices comprising the dorsal attention network" and with activity also in "posterior cingulate/precuneus . . . the superior parietal cortex (fronto-parietal network) and the occipital cortices (visual network). Resting state functional connectivity between bilateral prefrontal cortices encompassing the dorsal attention network and the right insula (salience network) was also associated with intelligence scores."
In humans, general intelligence is heritable and is dependent upon many genes interacting together (Bouchard, 2014). One can wonder whether some of these many genes determining intelligence might hold information about some of the environmental regularities in the world discussed above, including relational regularities such as causality and similarity. This is an area of research wide open for researchers in behavior genetics or cognitive genetics.
When considering human intelligence, the role of language is a significant factor. Much of human thinking involves the use of words. As Dicke and Roth (2016) state, "The evolution of a syntactical and grammatical language in humans most probably has served as an additional intelligence amplifier, which may have [also] happened in songbirds and psittacids" [parrots] as a consequence of convergent evolution--the evolution of similar characteristics in unrelated or distantly related species because of common selection pressures. As mentioned above, the precuneus of the parietal lobe may have played a significant role in the evolution of language by having involvement in the combination of ideas and concepts from different semantic domains (Rabini, et al., 2021).
Studies of brain damaged patients supply further insight into brain mechanisms involved in abstract concept formation and thinking dependent upon language. Patients with one type of frontotemporal dementia (FTD) are particularly impaired "in verbal concept formation (i.e. categorization based on abstract similarities between items). . . [with] especially the left frontal lobe, thought to be involved in abstract word processing (Lagarde, et al., 2015, p. 456).
Another Component of General Intelligence: Visual Imagery and Imagination
Another important component of general intelligence is ability to visualize--to form and manipulate visual images in imagination. Psychologists who study human intelligence using and analyzing intelligence tests label this the Broad Visual Perception factor (Gv), “which is an ability to generate, retain, retrieve and transform visual images” (Kvist and Gustafsson, 2008, p.422-423). This is one of the sub-factors of g (general intelligence) in Carroll’s (1993) widely accepted three-level model of human intelligence (see module on measuring human intelligence which follows). This visualization ability may have developed as an adaptation or perhaps as an "exaptation" (recruitment of an adaptation to a different function; see Gould and Vrba, 1982). On this view, portions of the visual system, including parts of visual, motor, and frontal and parietal association cortex, were recruited over evolutionary time to permit the formation and manipulation of visual images used to mentally test out probable effects of future behaviors before committing to them in the real physical world. Much of what we call "thought" is of this form (Koenigshofer, 2011, 2016, 2017). Note that some studies described above found activity in the cortical visual areas and frontal and parietal cortical areas to be associated with intelligence. Other studies, discussed above in the section on mental models, implicate the "default network" in both imagination and memory.

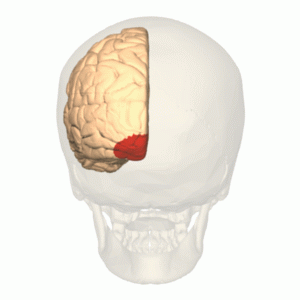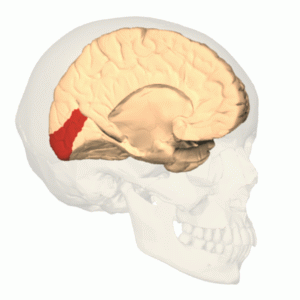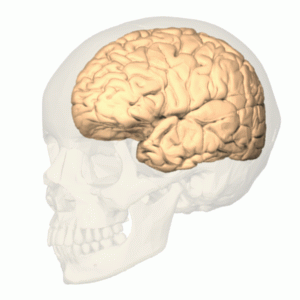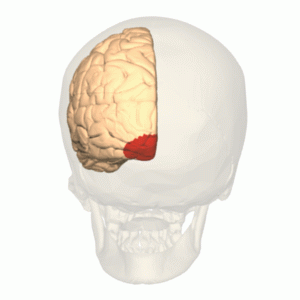

Figure \(\PageIndex{12}\): (Left) Left Parietal Lobe; (Center) Left Primary Visual Cortex; the Visual association cortex (not colored) immediately surrounds primary visual cortex; (Right) Left Primary Motor Cortex; premotor cortex (not colored) is located just anterior to primary motor cortex. (Images from Wikimedia Commons; https://commons.wikimedia.org/wiki/F..._animation.gif; https://commons.wikimedia.org/wiki/F..._animation.gif; https://commons.wikimedia.org/w/inde...=Go&type=image; by Polygon data were generated by Database Center for Life Science (DBCLS); licensed under the Creative Commons Attribution-Share Alike 2.1 Japan license; caption by Kenneth A. Koenigshofer, PhD).
There is experimental evidence for this claim. For example, brain imaging studies show that when humans imagine a particular movement, the same brain region that becomes active during the actual movement also becomes active during the imagined movement, even though no actual movement occurs. In both cases, the activated brain area shows nearly the same fMRI image (Ganis, et al., 2004; Kanwisher, 2009; Kosslyn, et al., 2006) suggesting that similar circuits are involved in imagining a movement and actually carrying out the movement. Similar results using fMRI have been reported for visual perception and mental imagery in the "mind's eye" with the greatest overlap in activation patterns for perception and imagery in imagination occurring in the frontal and parietal cortex (Ganis, et al., 2004). Mental testing of planned actions in the mind’s eye (guided by innate, implicit “knowledge” of cause-effect, predictive event covariation, and similarity relations) is highly adaptive because it is safe, fast, and efficient (Koenigshofer, 2017). Testing out possible behaviors and their likely effects in imagination can eliminate harmful or even fatal choices in the real world. In addition, mental testing of possible behavioral choices takes much less time and much less caloric expenditure than testing out behaviors by trial and error in the real physical world (think about mentally searching for your car keys until you find them mentally vs. actually moving to search your house, car, and garage to try to find them--the savings in time and in calories is clear). Because of these adaptive advantages, it is likely that there has been strong selection for neural mechanisms supporting visual imagination--a major component of human general intelligence and an integral part of what is generally referred to as "thinking" (Koenigshofer, 2011, 2016, 2017).
Experimental Support for Mental Imagery
Image Scanning
Seminal research in visual imagery was provided by Kosslyn's image-scanning experiments in the 1970s. Using the mental representation of a ship, subjects were instructed to shift their mental focus from one part of the ship to another. The reaction time of the subjects increased with distance between the two parts of the imagined ship, which indicates that we actually create a mental picture of scenes while trying to solve small cognitive tasks such as scanning the mental image of an object. Interestingly, Marmor and Zaback (1976) found that this visual ability can be observed also in people who are congenitally blind. Presuming, that the underlying processes are the same as in sighted subjects, it could be concluded that there is a deeper encoded system that has access to more than just the visual input.
Mental Rotation Task
Other advocates of visual spatial representation theory, Shepard and Metzler, developed the mental rotation task in 1971. Two spatially complex objects are presented to a participant at different angles and his/her job is to decide whether the objects are identical or not. The results show that the reaction times increases linearly with the rotation angle of the objects. The participants mentally rotate the objects in order to match the objects to one another. This method is called "mental chronometry".
This experiment was crucial for demonstrating the importance of imagery within cognitive psychology, because it showed the similarity of imagery to the processes of perception. For a mental rotation of 40° the subjects needed two seconds on average, whereas for a 140° rotation the reaction time increased to four seconds. Therefore, it can be concluded that people in general have a mental object rotation rate of 50° per second.
Spatial Frameworks
As mentioned above, researchers generally believe that mental models are perceptually based as indicated by fMRI studies which show extensive overlap between fMRI images of the perception of an object and its visualization. Indeed, people have been found to use spatial frameworks like those created for texts to retrieve spatial information about observed scenes (Bryant, 1991). Thus, people create the same sorts of spatial memory representations no matter if they read about an environment or see it themselves.
Size and the visual field
If an object is observed from different distances, it is harder to perceive details if the object is far away because the objects fill only a small part of the visual field. Kosslyn did an experiment in 1973 in which he wanted to find out if this is also true for mental images, to show the similarity of the spatial representation and the perception of real environment. He told participants to imagine objects which are far away and objects which are near. After asking the participants about details, he supposed that details can be observed better if the object is near and fills the visual field. He also told the participants to imagine animals with different sizes near by another. For example an elephant and a rabbit. The elephant filled much more of the visual field than the rabbit and it turned out that the participants were able to answer questions about the elephant more rapidly than about the rabbit. After that the participants had to imagine the small animal besides an even smaller animal, like a fly. This time, the rabbit filled the bigger part of the visual field and again, questions about the bigger animal were answered faster. The result of Kosslyn's experiments is that people can observe more details of an object if it fills a bigger part of their mental visual field. This provides evidence that mental images are represented visual-spatially.
Visual Imagery, Intelligence, and the Frontoparietal Network
Significantly, visual imagery in the mind’s eye can also be employed to mentally test novel combinations of causes and effects, similarity relations, and predictive relations (covariations) to discover new knowledge about hidden causes of events in the world, eventually leading to the creation of sophisticated scientific models of how the world works—human general intelligence at its best (Koenigshofer, 2017). For example, the famous theoretical physicist, Albert Einstein, used mental imagery extensively to do "thought experiments" that played a large role in his formulation of the theories of general and special relativity. Brain imaging studies using fMRI implicate parietal cortex in the use of imagination (Nair et al., 2003). Bruner (2010, p. S84) suggests that “the parietal lobe system ‘forms a neural image of surrounding space’ (Mountcastle, 1995 p. 389),” and perhaps of one’s potential future action in that space. Significantly, parietal cortex has strong linkages with prefrontal cortex forming a frontoparietal network: the inferior parietal lobule (further subdivided into the supramarginal gyrus, the temporoparietal junction, and the angular gyrus, see Figure 14.2.8) is primarily connected with dorsolateral prefrontal cortex (Bruner, 2010; see Figure 14.2.7), associated, in part, with abilities for abstract thought, while upper parietal regions, according to Bruner, are associated in the scientific literature with functions such as abstract representation, internal mental images, “imagined world[s],. . . and thought experiment” (i.e., imagination). Significantly, a number of other research groups using fMRI brain imaging techniques in humans also find evidence of a critical role of the frontoparietal network in human intelligence and general cognitive ability (Colom, et al., 2010; Jung & Haier, 2007; Sripada, et al., 2020; Vendetti & Bunge, 2014; Wendelken, et al., 2017; Yeo et al., 2016).
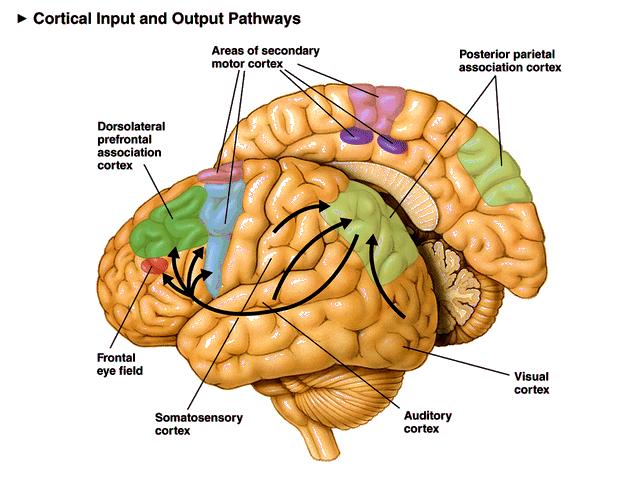
Figure \(\PageIndex{13}\): Connections of posterior parietal association cortex with dorsolateral prefrontal association cortex. (Image from Wikipedia; https://commons.wikimedia.org/wiki/F...ietal_Lobe.jpg; by Paskari; licensed under the Creative Commons Attribution-Share Alike 2.5 Generic, 2.0 Generic and 1.0 Generic license. Caption by Kenneth A. Koenigshofer, PhD.).
The emergence during human evolution of this visualization ability may help account for the development of modern human cognition, first appearing some 60,000-100,000 years ago (Mellars, 2005). Speculatively, superior abilities for imagination of the type described above might account, at least in part, for human competitive advantage over Neanderthals (Koenigshofer, 2017) perhaps contributing to Neanderthal extinction about 30,000 years ago (Watson and Berry, 2009). Consistent with this view, morphological studies suggest enhanced parietal lobe development in modern humans compared to Neanderthals (Bruner, 2010); by contrast, recent studies show little relative enlargement of the frontal lobes in humans compared to apes (Barton and Venditti, 2013). At least one post-mortem anatomical study of Einstein's brain suggested unusual enlargement of the inferior parietal lobules in parietal lobes along with a number of other atypical features in the occipital lobes, and throughout the cerebral cortex (Chen et al., 2014; see Carrillo-Mora et al., 2015). Another study of Einstein's brain by Falk et al. (2013, p. 1304) reports "Einstein’s brain has an extraordinary prefrontal cortex, which may have contributed to the neurological substrates for some of his remarkable cognitive abilities. The primary somatosensory and motor cortices near the regions that typically represent face and tongue are greatly expanded in the left hemisphere. Einstein’s parietal lobes are also unusual and may have provided some of the neurological underpinnings for his visuospatial and mathematical skills, as others have hypothesized. Einstein’s brain has typical frontal and occipital shape asymmetries (petalias) and grossly asymmetrical inferior and superior parietal lobules" with relative enlargement of the left inferior parietal module. Chen et al., referred to above, found exceptional enlargement of both inferior parietal lobules (which include the supramarginal gyrus and angular gyrus (Figure 14.2.8), involved in multiple functions including number processing, "mentalizing," and spatial cognition) in Einstein's brain compared to controls. Einstein was famous for his use of visualization to explore his ideas in theoretical physics through "thought experiments" in his "mind's eye." Additional evidence regarding Einstein's exceptional parietal lobes was provided by studies of cortical glial cells. A study by Diamond et al. (1985) of Einstein's brain found that the only cortical area of the four cortical areas examined with a greater number of glial cells per neuron, compared to controls, was the left posterior parietal cortex, a finding interpreted by the authors to mean that this area of Einstein's brain must have been exceptionally active during his lifetime.
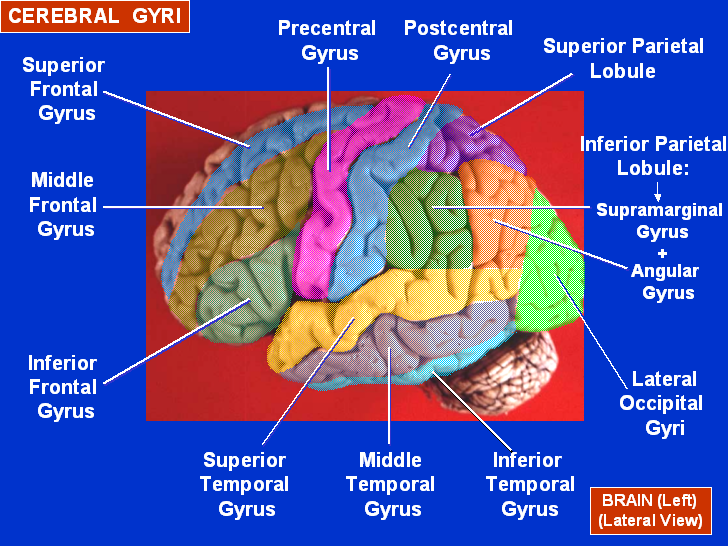
Figure \(\PageIndex{14}\): Cortical gyri, including the left inferior parietal lobule composed of the supramarginal and angular gyri. (Image from Wikimedia Commons; https://commons.wikimedia.org/wiki/F...al_Surface.png; by John A. Beal, Ph.D.; licensed under the Creative Commons Attribution 2.5 Generic license.
This capacity for visualization permitting mental trial and error likely occurs to some degree in non-human animal species as well. Kohler's (1925/1959) classic work showing the use of "insight" by chimpanzees in problem solving offers evidence for this hypothesis, assuming that insight involves mental trial and error. Additional evidence suggesting mental trial and error in imagination in a variety of animals is provided by experiments showing insight in some orangutans, gorillas, and chimpanzees (Mendes, et al., 2007; Hanus, et al., 2011) and in rooks--birds in the same Corvid family as ravens and crows (Bird & Emery, 2009a, 2009b).
One key feature of more traditional approaches to the study of human intelligence is that they emphasize aspects of human intelligence and thinking, such as language and computation, that are especially important for life in complex modern society. As described in the next module, some psychologists have hypothesized multiple kinds of human intelligence. Others focus primarily on how to measure intelligence, and still others study individual differences in intelligence among people. Because these psychologists tend to be interested in practical application of their tests in school and work settings, they tend to give relatively little attention to theories about the evolutionary origins or biological functions of intelligence. Nevertheless, intelligence tests are very useful predictors of success in environments such as school, military, and work settings. All involve measurement of mental skills important to successful adaptation to the demands of the physical and social environments typical of modern, industrialized societies.
Summary
Intelligence and cognition are complex psychological adaptations involving many different areas of the brain interacting together. Intelligence in the broadest sense involves the formation of sophisticated mental models of the world and how it works. These models include representations of things, events, and the predictive, similarity, and causal relations among them, as well as representations of space and time. Mental models of the physical and social environments permit prediction of future events, providing powerful evolutionary advantage. Mental models include the formation of categories, based on similarities, which allow inferences based on assignment of newly encountered things and events to previously formed categories. Mental models exploit the innate disposition to understand cause-effect relations in the environment, leading to knowledge about what causes what. This knowledge allows manipulation of causes to affect future environmental outcomes toward better adaptation. General intelligence emerges during evolution from genetic internalization of universal, abstract relational features of the world including cause-effect, event covariation, and similarity, each of which plays a role in thought and reasoning about the environment (Koenigshofer, 2017). Spearman (1904, 1925) discovered and was first to describe general intelligence, which he believed consisted of ability to recognize "relations and correlates" in the environment. Another aspect of general intelligence is ability to visualize, to form and manipulate visual images in imagination. This ability may have developed as an exaptation or recruitment of portions of the visual and motor systems, including parietal association cortex. This would allow some visual and motor circuits to form internally generated mental images that can be mentally manipulated to test out probable effects of possible future behaviors before committing to them in the physical world. Because this ability to visualize outcomes in the "mind's eye" is much safer and saves time and calories compared to testing outcomes in actual physical behavior, it is likely that there has been strong selection pressure for ability to manipulate mental images to plan future action. This component of intelligence provides a powerful mechanism for maximizing the adaptive outcomes of behavior and therefore provides enormous selective advantage--a strong impetus for its evolution as a central feature of intelligence (Koenigshofer, 2017).
References
Adams, S. S., & Burbeck, S. (2012). Beyond the octopus: From general intelligence toward a human-like mind. In Theoretical foundations of artificial general intelligence (pp. 49-65). Atlantis Press, Paris.
Amodio, P. (2019). Octopus intelligence: the importance of being agnostic. Animal Sentience 26(1). 272
Anderson, J. R. (2009). How can the human mind occur in the physical universe?. Oxford University Press.
Baron-Cohen, S., Wheelwright, S., Spong, A., Scahill, V., & Lawson, J. (2001). Are intuitive physics and intuitive psychology independent? A test with children with Asperger Syndrome. Journal of developmental and learning disorders, 5 (1), 47-78.
Barton, R. A., and Venditti, C. (2013). Human frontal lobes are not relatively large. Proceedings of the National Academy of Sciences, 110 (22), 9001-9006.
Behrens, T. E., Muller, T. H., Whittington, J. C., Mark, S., Baram, A. B., Stachenfeld, K. L., & Kurth-Nelson, Z. (2018). What is a cognitive map? Organizing knowledge for flexible behavior. Neuron, 100(2), 490-509.
Bird, C. D., & Emery, N. J. (2009a). Rooks use stones to raise the water level to reach a floating worm. Current Biology, 19 (16), 1410-1414.
Bird, C. D., & Emery, N. J. (2009b). Insightful problem solving and creative tool modification by captive nontool-using rooks. Proceedings of the National Academy of Sciences, 106 (25), 10370-10375.
Blakemore, S., Fonlupt, P., Pachot-Clouard, M., Darmon, C., Boyer, P., Meltzoff, A., Segebarth, C., Decety, J. (2001). How the brain perceives causality: an event-related fMRI study, NeuroReport 12: 3741 – 3746.
Bouchard, T. J. (2014). Genes, evolution and intelligence. Behavior genetics, 44(6), 549-577.
Broschard, M. B., Kim, J., Love, B. C., Wasserman, E. A., & Freeman, J. H. (2021). Prelimbic cortex maintains attention to category-relevant information and flexibly updates category representations. Neurobiology of Learning and Memory, 107524
Bruner, E. (2010). Morphological differences in the parietal lobes within the human genus. Current Anthropology, 51 (S1), S77-S88
Clark, A. (2013). Whatever next? Predictive brains, situated agents, and the future of cognitive science. Behavioral and Brain Sciences, 36(3), 181–204.
Clark, A. (2016). Surfing uncertainty: Prediction, action, and the embodied mind. New York: Oxford University Press.
Carrillo-Mora, P., Magaña-Vázquez, K., May-López, S. S., & Mondragón-Ramírez, N. A. (2015). What did Einstein have that I don’t? Studies on Albert Einstein’s brain. Neurosciences and History; 3(3): 125-129; https://nah.sen.es/vmfiles/abstract/...5125_129EN.pdf; retrieved 12/23/21.
Carroll, J. B. (1993). Human cognitive abilities: A survey of factor-analytic studies (No. 1). Cambridge University Press.
Cattell, R. B. (1987). Intelligence: Its structure, growth and action. New York: Elsevier Science.
Chen, H., Chen, S., Zeng, L., Zhou, L., & Hou, S. (2014). Revisiting Einstein's brain in Brain Awareness Week. Bioscience trends, 8 (5), 286-289.
Churchland, P. M. (1985, 2013). Matter and consciousness. MIT press. Cambridge, Mass.
Colom, R., Karama, S., Jung, R. E., & Haier, R. J. (2010). Human intelligence and brain networks. Dialogues in clinical neuroscience, 12 (4), 489.
Cosmides, L., & Tooby, J. (1992). Cognitive adaptations for social exchange. In J. H. Barkow, L. Cosmides, & J. Tooby (Eds.), The adapted mind: Evolutionary psychology and the generation of culture (pp. 163–228). New York: Oxford University Press.
Cosmides, L., & Tooby, J. (2003). “Evolutionary Psychology: Theoretical Foundations.” In: Encyclopedia of Cognitive Science. Ed. Lynn Nadel. London: Macmillan, 54–64.
Cosmides, L., & Tooby, J. (2007). Evolutionary psychology: A primer. https://www.cep.ucsb.edu/primer.html; retrieved 12/23/2021.
Dawkins, Richard (1976). The Selfish Gene. Best Books.
DeLoache, J. S., & LoBue, V. (2009). The narrow fellow in the grass: Human infants associate snakes and fear. Developmental science, 12 (1), 201-207.
De Pasquale, R., Beckhauser, T. F., Hernandes, M. S., & Britto, L. R. G. (2014). LTP and LTD in the visual cortex require the activation of NOX2. Journal of Neuroscience, 34 (38), 12778-12787.
D'Esposito, M., & Postle, B. R. (2015). The cognitive neuroscience of working memory. Annual review of psychology, 66, 115-142.
Diamond MC, Scheibel AB, Murphy GM Jr, Harvey T. On the brain of a scientist: Albert Einstein. Exp Neurol. 1985;88:198-204.
Dicke U, Roth G. (2016). Neuronal factors determining high intelligence. Phil. Trans. R. Soc. B 371: 20150180. http://dx.doi.org/10.1098/rstb.2015.0180; retrieved September 14, 2021.
Daw, N., Rao, Y., Wang, X. F., Fischer, Q., & Yang, Y. (2004). LTP and LTD vary with layer in rodent visual cortex. Vision research, 44(28), 3377-3380.
Durham, W. H. (1991). Coevolution: Genes, culture and human diversity. Palo Alto, CA: Stanford University Press.
Ellis, B. J., & Ketelaar, T. (2002). Commentary: Clarifying the Foundations of Evolutionary Psychology: A Reply to Lloyd and Feldman. Psychological Inquiry, 13 (2), 157-164.
Emery, N. J., and Clayton, N. S. (2004). The mentality of crows: convergent evolution of intelligence in corvids and apes. Science, 306 (5703), 1903-1907.
Falk, D., Lepore, F. E., & Noe, A. (2013). The cerebral cortex of Albert Einstein: a description and preliminary analysis of unpublished photographs. Brain, 136 (4), 1304-1327.
Fonlupt, P. (2003). Perception and judgment of physical causality involve different brain structures, Cogn. Brain Res. 17, 248 – 254.
Frith, C. (2007). Making Up the Mind: How the Brain Creates Our Mental World. Oxford: Blackwell.
Frith, U. & Frith, C. D. (2010). The Social Brain: Allowing Humans to Boldly Go Where No Other Species Has Been. Philosophical Transactions of the Royal Society B Biological Sciences 365(1537): 165–76
Fugelsang, J. A., & Dunbar, K. N. (2005). Brain-based mechanisms underlying complex causal thinking. Neuropsychologia, 43 (8), 1204-1213.
Fugelsang, J. A., Roser, M. E., Corballis, P. M., Gazzaniga, M. S., & Dunbar, K. N. (2005). Brain mechanisms underlying perceptual causality. Cognitive brain research, 24 (1), 41-47.
Gallistel C.R. (1995). The replacement of general purpose theories with adaptive specializations. In M.S. Gazzaniga (Ed.), The Cognitive Neurosciences (pp. 1255–67). Cambridge, MA: MIT Press.
Gangestad, S. W., & Simpson, J. A. (2000). The evolution of human mating: Trade-offs and strategic pluralism. Behavioral and Brain Sciences, 23, 573–644.
Ganis, G., Thompson, W. L., & Kosslyn, S. M. (2004). Brain areas underlying visual mental imagery and visual perception: an fMRI study. Cognitive Brain Research, 20(2), 226-241.
Geertz, A. W. (2016). The meaningful brain. In: Mental Culture: Classical Social Theory and the Cognitive Science of Religion. Edited by Dimitris Xygalatas and William W. McCorkle Jr., 176-196.
Goldman-Rakic, P. S. (1996). The prefrontal landscape: implications of functional architecture for understanding human mentation and the central executive. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences, 351(1346), 1445-1453.
Goodall, J. (2000). In the shadow of man (Vol. 4113). Houghton Mifflin Harcourt.
Gopnik, A. (2010). How babies think. Scientific American, 303 (1), 76-81.
Gopnik, A. (2012). Scientific thinking in young children: Theoretical advances, empirical research, and policy implications. Science, 337 (6102), 1623-1627.
Gopnik, A., and Sobel, D. M. (2000). Detecting blickets: How young children use information about novel causal powers in categorization and induction. Child development, 71 (5),1205-1222.
Gopnik, A., Sobel, D. M., Schulz, L. E., and Glymour, C. (2001). Causal learning mechanisms in very young children: two-, three-, and four-year-olds infer causal relations from patterns of variation and covariation. Developmental psychology, 37 (5), 620.
Gopnik, A., & Wellman, H. M. (2012). Reconstructing constructivism: Causal models, Bayesian learning mechanisms, and the theory theory. Psychological bulletin, 138 (6), 1085.
Gopnik, A., Glymour, C., Sobel, D. M., Schulz, L. E., Kushnir, T., & Danks, D. (2004). A theory of causal learning in children: Causal maps and Bayes nets. Psychological Review, 111, 3–32.
Gould, S. J., & Vrba, E. S. (1982). Exaptation—a missing term in the science of form. Paleobiology, 8 (1), 4-15.
Haier, R. J., Siegel, B., Tang, C., Abel, L., & Buchsbaum, M. S. (1992). Intelligence and changes in regional cerebral glucose metabolic rate following learning. Intelligence, 16 (3-4), 415-426.
Hanus, D., Mendes, N., Tennie, C., & Call, J. (2011). Comparing the performances of apes (Gorilla gorilla, Pan troglodytes, Pongo pygmaeus) and human children (Homo sapiens) in the floating peanut task. PloS one, 6 (6), e19555.
Hawkins, J., Ahmad, S., & Cui, Y. (2017). A theory of how columns in the neocortex enable learning the structure of the world. Frontiers in neural circuits, 81.
Hayashi, M. J., Kantele, M., Walsh, V., Carlson, S., & Kanai, R. (2014). Dissociable neuroanatomical correlates of subsecond and suprasecond time perception. Journal of Cognitive Neuroscience, 26(8), 1685-1693.
Hearne, L. J., Mattingley, J. B., & Cocchi, L. (2016). Functional brain networks related to individual differences in human intelligence at rest. Scientific reports, 6 (1), 1-8.
Huber, L., & Gajdon, G. K. (2006). Technical intelligence in animals: the kea model. Animal cognition, 9 (4), 295-305.
Hume, D. (1748/1988). An Enquiry Concerning Human Understanding. La Salle, IL: Open Court (Originally published 1748).
Ivry, R. B., & Keele, S. W. (1989). Timing functions of the cerebellum. Journal of cognitive neuroscience, 1 (2), 136-152.
Ivry, R. B., Spencer, R. M., Zelaznik, H. N., & Diedrichsen, J. (2002). The cerebellum and event timing. Annals of the new York Academy of Sciences, 978 (1), 302-317.
Ivry, R. B., & Spencer, R. M. (2004). Evaluating the role of the cerebellum in temporal processing: beware of the null hypothesis. Brain, 127 (8), e13-e13.
Jung, R. E., & Haier, R. J. (2007). The Parieto-Frontal Integration Theory (P-FIT) of intelligence: converging neuroimaging evidence. Behavioral and Brain Sciences, 30 (2), 135-154.
Kanwisher, N. (2009, May). The Specialized Brain. Keynote Address. Annual Meeting of the Association for Psychological Science. San Francisco.
Kaufman, S. B., DeYoung, C. G., Reis, D. L., and Gray, J. R. (2011). General intelligence predicts reasoning ability even for evolutionarily familiar content. Intelligence, 39(5), 311–322.
Khemlani, S. S., Barbey, A. K., & Johnson-Laird, P. N. (2014). Causal reasoning with mental models. Frontiers in human neuroscience, 8, 849.
Koenigshofer, K. A. (2011). Mind Design: The Adaptive Organization of Human Nature, Minds, and Behavior. Pearson Education, Boston.
Koenigshofer, K. A. (2016). Mind Design: The Adaptive Organization of Human Nature, Minds, and Behavior, Revised Edition. Amazon e-book.
Koenigshofer, K. A. (2017). General Intelligence: Adaptation to Evolutionarily Familiar Abstract Relational Invariants, Not to Environmental or Evolutionary Novelty. The Journal of Mind and Behavior, 38 (2), 119-153.
Köhler, W. (1925/1959). The mentality of apes (E. Winter, Trans.). New York, USA: Vintage Books
Kosslyn, S. M., Thompson, W. L., & Ganis, G. (2006). The Case for Mental Imagery. Oxford University Press
Kubricht, J. R., Holyoak, K. J., & Lu, H. (2017). Intuitive physics: Current research and controversies. Trends in cognitive sciences, 21(10), 749-759.
Kvist, A. V., and Gustafsson, J. E. (2008). The relation between fluid intelligence and the general factor as a function of cultural background: A test of Cattell's investment theory. Intelligence, 36 (5), 422-436.
Lagarde, J., Valabregue, R., Corvol, J. C., Garcin, B., Volle, E., Le Ber, I., ... & Levy, R. (2015). Why do patients with neurodegenerative frontal syndrome fail to answer:‘In what way are an orange and a banana alike?’. Brain, 138(2), 456-471.
Lara, A. H., & Wallis, J. D. (2015). The role of prefrontal cortex in working memory: a mini review. Frontiers in systems neuroscience, 9, 173.
Leslie, A. M., & Keeble, S. (1987). Do six-month-old infants perceive causality?. Cognition, 25(3), 265-288.
Lloyd, E. A., & Feldman, M. W. (2002). Commentary: Evolutionary psychology: A view from evolutionary biology. Psychological Inquiry, 13 (2), 150-156.
Mather, J. (2019). What is in an octopus’s mind? Animal Sentience 26 (1).
Mather, J. A., and Dickel, L. (2017). Cephalopod complex cognition. Current Opinion in Behavioral Science 16, 131–137. doi:10.1016/J.COBEHA.2017.06.008.
Mellars, P. (2005). The impossible coincidence. A single‐species model for the origins of modern human behavior in Europe. Evolutionary Anthropology: Issues, News, and Reviews, 14 (1), 12-27.
Mendes, N., Hanus, D., & Call, J. (2007). Raising the level: Orangutans use water as a tool. Biology Letters, 3, 453– 455. doi:10.1098/rsbl.2007.0198
Miller, E. K., & Cohen, J. D. (2001). An integrative theory of prefrontal cortex function. Annual review of neuroscience, 24 (1), 167-202.
Nair, D. G., Purcott, K. L., Fuchs, A., Steinberg, F., and Kelso, J. S. (2003). Cortical and cerebellar activity of the human brain during imagined and executed unimanual and bimanual action sequences: a functional MRI study. Cognitive brain research, 15 (3), 250-260.
Nave K, Deane G, Miller M, Clark A. (2020). Wilding the predictive brain. WIREs Cogn Sci. 2020;11:e1542. https://doi.org/10.1002/wcs.1542
Numan, R. (2015). A prefrontal-hippocampal comparator for goal-directed behavior: the intentional self and episodic memory. Frontiers in behavioral neuroscience, 9, 323.
Oakes, L. M. (1994). Development of infants' use of continuity cues in their perception of causality. Developmental Psychology, 30 (6), 869.
Oishi, N., Nomoto, M., Ohkawa, N., Saitoh, Y., Sano, Y., Tsujimura, S., ... & Inokuchi, K. (2019). Artificial association of memory events by optogenetic stimulation of hippocampal CA3 cell ensembles. Molecular brain, 12 (1), 1-10.
Panksepp, J., & Panksepp, J. B. (2000). The seven sins of evolutionary psychology. Evolution and cognition, 6 (2), 108-131.
Penn, D. C., and Povinelli, D. J. (2007). Causal Cognition in Human and Nonhuman Animals: A Comparative, Critical Review. Annual Review of Psychology 58, 97–118. doi:10.1146/annurev.psych.58.110405.085555.
Penn, D. C., Holyoak, K. J., and Povinelli, D. J. (2008). Darwin's mistake: Explaining the discontinuity between human and nonhuman minds. Behavioral and Brain Sciences, 31 (02), 109-130.
Pinker, S. (2007). The stuff of thought: Language as a window into human nature. New York: Penguin Books.
Rabini, G., Ubaldi, S., & Fairhall, S. L. (2021). Combining concepts across categorical domains: a linking role of the precuneus. Neurobiology of Language, 1-50.
Reader, S. M., Hager, Y., and Laland, K. N. (2011). The evolution of primate general and cultural intelligence. Philosophical Transactions of the Royal Society B: Biological Sciences, 366(1567), 1017–1027. doi: 10.1098/rstb.2010.0342
Roser, M. E., Fugelsang, J. A., Dunbar, K. N., Corballis, P. M., & Gazzaniga, M. S. (2005). Dissociating processes supporting causal perception and causal inference in the brain. Neuropsychology, 19 (5), 591.
Schacter, D. L., Addis, D. R., Hassabis, D., Martin, V. C., Spreng, R. N., & Szpunar, K. K. (2012). The future of memory: remembering, imagining, and the brain. Neuron, 76 (4), 677-694.
Schafer, M., & Schiller, D. (2020). The Brain's Social Road Maps. Scientific American, 322 (2), 31-35.
Scholl, B. J., & Nakayama, K. (2002). Causal capture: Contextual effects on the perception of collision events. Psychological science, 13 (6), 493-498.
Soto, F. A., & Wasserman, E. A. (2012). Visual object categorization in birds and primates: Integrating behavioral, neurobiological, and computational evidence within a “general process” framework. Cognitive, Affective, & Behavioral Neuroscience, 12 (1), 220-240.
Spearman, C. (1904). "General Intelligence," objectively determined and measured. The American Journal of Psychology, 15(2), 201-292.
Spearman, C. (1925). Some Issues in the Theory of`'g' (including the Law of Diminishing Returns). Nature, 116, 436-439.
Shepard, R.N. (1987a). Evolution of a mesh between principles of the mind and regularities of the world. In J. Dupre ( Ed.), The Latest and the Best: Essays on Evolution and Optimality. Cambridge, MA: MIT Press.
Shepard, R.N. (1987b). Toward a universal law of generalization for psychological science. Science, 237, 1317-1323.
Shepard, R.N. (1992). The perceptual organization of colors: an adaptation to regularities of the terrestrial world? In J.H. Barkow, L. Cosmides, and J. Tooby (Eds.), The Adapted Mind: Evolutionary Psychology and the Generation of Culture. New York: Oxford University Press.
Shepard, R.N. (1994). Perceptual-cognitive universals as reflections of the world. Psychonomic Bulletin and Review 1:2-28. .
Shepard, R. N. (2001). Perceptual-cognitive universals as reflections of the world. Behavioral and brain sciences, 24 (04), 581-601.
Siegel, M., Buschman, T. J., and Miller, E. K. (2015). Cortical information flow during flexible sensorimotor decisions. Science 348, 1352–1355. doi: 10.1126/science.aab0551
Spreng, R. N., Gerlach, K. D., Turner, G. R., & Schacter, D. L. (2015). Autobiographical planning and the brain: Activation and its modulation by qualitative features. Journal of cognitive neuroscience, 27 (11), 2147-2157.
Sripada, C., Angstadt, M., Rutherford, S., Taxali, A., & Shedden, K. (2020). Toward a “treadmill test” for cognition: Improved prediction of general cognitive ability from the task activated brain. Human brain mapping, 41(12), 3186-3197.
Striedter, G. F. (2005). Principles of brain evolution. Sinauer associates.
Thagard, P. (2010). How brains make mental models. In Model-based reasoning in science and technology (pp. 447-461). Springer, Berlin, Heidelberg.
Tolman, E. C. (1948). Cognitive maps in rats and men. Psychological review, 55 (4), 189.
Tooby, J., & Cosmides, L. (2015). The theoretical foundations of evolutionary psychology. The handbook of evolutionary psychology: Foundations (2nd ed., Vol. 1, pp. 3–87). Hoboken, NJ: John Wiley.
Van Den Heuvel, M. P., Stam, C. J., Kahn, R. S., & Pol, H. E. H. (2009). Efficiency of functional brain networks and intellectual performance. Journal of Neuroscience, 29 (23), 7619-7624.
Vendetti, M. S., & Bunge, S. A. (2014). Evolutionary and developmental changes in the lateral frontoparietal network: a little goes a long way for higher-level cognition. Neuron, 84 (5), 906-917.
Waltz, J. A., Knowlton, B. J., Holyoak, K. J., Boone, K. B., Mishkin, F. S., de Menezes Santos, M., ... & Miller, B. L. (1999). A system for relational reasoning in human prefrontal cortex. Psychological science, 10 (2), 119-125.
Wasserman, E. A., Wasserman, E. A., & Zentall, T. R. (Eds.). (2006). Comparative cognition: Experimental explorations of animal intelligence. Oxford University Press, USA.
Watson, J. D., and Berry, A. (2009). DNA: The secret of life. New York: Knopf.
Wendelken, C., Ferrer, E., Ghetti, S., Bailey, S. K., Cutting, L., & Bunge, S. A. (2017). Frontoparietal structural connectivity in childhood predicts development of functional connectivity and reasoning ability: A large-scale longitudinal investigation. Journal of Neuroscience, 37 (35), 8549-8558.
Yeo, R. A., Ryman, S. G., Pommy, J., Thoma, R. J., & Jung, R. E. (2016). General cognitive ability and fluctuating asymmetry of brain surface area. Intelligence, 56, 93-98.
Attributions
Section 14.2, "Intelligence, Cognition, and Language as Psychological Adaptations," is original material written by Kenneth A. Koenigshofer, PhD, licensed under CC BY 4.0, with the exception of the section titled "Experimental Evidence for Mental Imagery" which is adapted by Kenneth A. Koenigshofer, Ph.D., from Cognitive Psychology and Cognitive Neuroscience, Wikibooks, https://en.wikibooks.org/wiki/Cognit...cience/Imagery; text is available under the Creative Commons Attribution-ShareAlike License
Images from Wikimedia Commons.