
8.1.3: The Origin of Primates
- Last updated
- Save as PDF
- Page ID
- 136421
Paleocene: Mammals in the Wake of Dinosaur Extinctions
Placental mammals, including primates, originated in the Mesozoic Era (approximately 251 million to 65.5 million years ago), the Age of Dinosaurs. During this time, most placental mammals were small, probably nocturnal, and probably avoided predators via camouflage and slow, quiet movement. It has been suggested that the success and diversity of the dinosaurs constituted a kind of ecological barrier to Mesozoic mammals. The extinction of the dinosaurs (and many other organisms) at the end of the Cretaceous Period (approximately 145.5–65.5 million years ago) might have opened up these ecological niches, leading to the increased diversity and disparity in mammals of the Tertiary Period (approximately 65.5–2.5 million years ago).
The Paleocene was the first epoch in the Age of Mammals. Soon after the Cretaceous-Tertiary (K-T) extinction event, new groups of placental mammals appear in the fossil record. Many of these groups achieved a broad range of sizes and lifestyles as well as a great number of species before declining sometime in the Eocene (or soon thereafter). These groups were ultimately replaced by the modern orders of placental mammals (Figure 8.3). It is unknown whether these replacements occurred gradually, for example by competitive exclusion, or rapidly, perhaps by sudden geographic dispersals with replacement. In some senses, the Paleocene might have been a time of recovery from the extinction event; it was cooler and more seasonal globally than the subsequent Eocene.
 Figure \(\PageIndex{1}\): Depiction of Eocene flora and fauna in North America.
Figure \(\PageIndex{1}\): Depiction of Eocene flora and fauna in North America.Plesiadapiforms, the Archaic Primates
The Paleocene epoch saw the emergence of several families of mammals that have been implicated in the origin of primates. These are the plesiadapiforms. Plesiadapiforms are archaic primates, meaning that they possessed some primate features and lacked others. The word plesiadapiform means “almost adapiform,” a reference to some similarities between some plesiadapiforms and some adapiforms (or adapoids; later-appearing true primates)—mainly in the molar teeth. Because enamel fossilizes better than other parts of the body, the molar teeth are the parts most often found and first discovered for any new species. Thus, dental similarities were often the first to be noticed by early mammalian paleontologists, partly explaining why plesiadapiforms were thought to be primates. Major morphological differences between plesidapiforms and euprimates (true primates) were observed later when more parts of plesiadapiform skeletons were discovered. Many plesiadapiforms have unusual anterior teeth and most have digits possessing claws rather than nails. So far, no plesiadapiform ever discovered has a postorbital bar (seen in extant strepsirrhines) or septum (as seen in haplorhines), and whether or not the auditory bulla was formed by the petrous bone remains unclear for many plesiadapiform specimens. Nevertheless, there are compelling reasons (partly from new skeletal material) for including plesidapiforms within the Order Primates.
Geographic and Temporal Distribution
Purgatorius is generally considered to be the earliest primate. This Paleocene mammal is known from teeth that are very primitive for a primate. It has some characteristics that suggest it is a basal plesiadapiform, but there is very little to link it specifically with euprimates (see Clemens 2004). Its ankle bones suggest a high degree of mobility, signaling an arboreal lifestyle (Chester et al. 2015). Purgatorius is primitive enough to have given rise to all primates, including the plesiadapiforms. Plesiadapiform families were numerous and diverse during parts of the Paleocene in western North America and western Europe, with some genera (e.g., Plesiadapis; see Table 8.1) living on both continents (Figure 8.4). Thus, there were probably corridors for plesiadapiform dispersal between the two continents, and it stands to reason that these mammals were living all across North America, including in the eastern half of the continent and at high latitudes. A few plesiadapiforms have been described from Asia (e.g., Carpocristes), but the affinities of these remain uncertain.
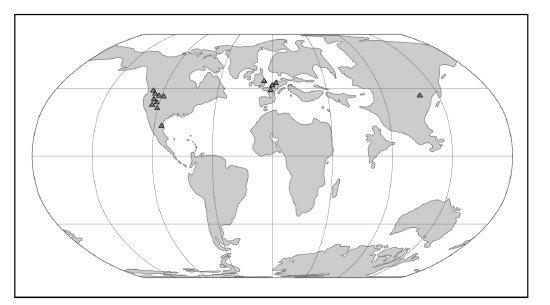 Figure \(\PageIndex{2}\): Map of the world in the Paleocene, highlighting plesiadapiform localities.
Figure \(\PageIndex{2}\): Map of the world in the Paleocene, highlighting plesiadapiform localities.General Morphological Features
Although there is much morphological variation among the families of plesiadapiforms, there are some common features of the group. Most plesiadapiforms were small, the largest being about three kilograms (approximately 7 lbs.; Plesiadapis cookei). They had small brains and fairly large snouts, with variable eye size (as deduced from the bony orbits). In general, the eyes faced more laterally than in euprimates. Most plesiadapiforms have large incisors relative to the molars and in some species, the lower incisors (usually one pair) are reminiscent of long daggers or spears. In many plesiadapiforms, the upper central incisors are also very unusual, with small cuspules spaced out like fingers, having some unknown function perhaps related to seizing or cropping food. Many species show reduction and/or loss of the canine and anterior premolars, with the resulting formation of a rodent-like diastema; this probably implies a herbivorous diet. The spaces available for the chewing musculature are very large, and most plesiadapiforms probably had very powerful chewing muscles, perhaps capable of processing very tough foods (e.g., leaves). Some families appear to have had very specialized diets, as suggested by unusual tooth and jaw shapes. For example, an enlarged, laterally compressed, blade-like lower premolar appears to have evolved via convergent evolution in two different families, the Carpolestidae and the Saxonellidae.
Arguably the most interesting and unusual family of plesiadapiforms is the Carpolestidae. This family contains three major genera and a few minor ones. They are almost exclusively from North America (with a couple of possible members from Asia), and mainly from the Middle and Late Paleocene. Their molars are not very remarkable, being quite similar to those of some other plesiadapiforms (e.g., Plesiadapidae). However, nearly everything else is unusual. Their lower posterior premolars (p4) are laterally compressed and blade-like with vertical serrations topped by tiny cuspules. This unusual dental morphology is termed “plagiaulacoid” (Simpson 1933). It is similar to the condition in some living and fossil marsupials, but in marsupials, the blade-like lower tooth slides across a similar-looking blade-like upper tooth. In carpolestids, the blade-like tooth meets upper premolars that look completely different. The upper premolar occlusal surfaces are broad and are covered with many small cuspules; the blade-like lower premolar might have cut across these cuspules, between them, or both.
 Figure \(\PageIndex{3}\): An artistic rendition of Carpolestes simpsoni moving along a small diameter support.
Figure \(\PageIndex{3}\): An artistic rendition of Carpolestes simpsoni moving along a small diameter support.Many plesiadapiforms have hallmarks of arboreality in the skeleton, though the long bones are rather robust. Instead of having nails, most taxa had sharp claws on most or all of the digits. The extremities show signs that at least some of these archaic primates had grasping abilities comparable to those of primates and some arboreal marsupials. Nearly complete skeletons are known for several North American plesiadapiforms, and these have yielded a tremendous wealth of information on locomotor and foraging habits. Many plesiadapiforms appear to have been well adapted to clinging to vertical substrates (like a broad tree trunk) using their sharp claws, propelling themselves upward using powerful hindlimbs, bounding along horizontal supports, grasping smaller branches, and moving head-first down tree trunks. In carpolestids in particular, the skeleton appears to have been especially well adapted to moving slowly and carefully in small terminal branches. The big toe likely was especially good at grasping (Figure 8.5). There is a single specimen of a flattened nail-bearing distal big toe bone of Carpolestes simpsoni (Table 8.1), and this feature suggests affinities with euprimates.
Debate: Relationship of Plesiadapiforms to True Primates
In the middle of the 20th century, treeshrews (Order Scandentia) were often considered part of the Order Primates, based on anatomical similarities between some treeshrews and primates. For many people, plesiadapiforms represented intermediates between primates and treeshrews, so plesiadapiforms were included in Primates as well.
Later, studies of reproduction and brain anatomy in treeshrews and lemurs suggested that treeshrews are not primates (e.g., Martin 1968). This was soon followed by the suggestion to also expel plesiadapiforms (Martin 1972). Like treeshrews, plesiadapiforms lack a postorbital bar, nails, and details of the ear region that characterize true primates. Many paleoanthropologists were reluctant to accept this move to banish plesiadapiforms (e.g. P. D. Gingerich 1980).
Later, Beard (1990) found that in some ways, the digits of paromomyid plesiadapiforms are actually more similar to those of dermopterans (Order Dermoptera), the closest living relatives of primates, than they are to those of primates themselves (but see Krause 1991). At the same time, Kay and colleagues (1990) found that cranial circulation patterns and auditory bulla morphology in the paromomyid, Ignacius (Table 8.1), are more like those of dermopterans than like those of primates.
For many anthropologists, this one-two punch effectively removed plesiadapiforms from the Order Primates. In the last two decades, the tide of opinion has turned again, with many researchers including plesiadapiforms in the Order Primates. New and more complete specimens demonstrate that the postcranial skeletons of plesiadapiforms, including the hands and feet, were primate-like, not dermorpteran-like (Bloch and Boyer 2002, 2007). New fine-grained CT scans of relatively complete plesiadapiform skulls revealed that they share some key traits with primates to the exclusion of other placental mammals (Bloch and Silcox 2006). Most significant was the suggestion that Carpolestes simpsoni possessed an auditory bulla formed by the petrosal bone, like in all living primates.
The debate about the status of plesiadapiforms continues, owing to a persistent lack of key bones in some species and owing to genuine complexity of the anatomical traits involved. Maybe plesiadapiforms were the primitive stock from which all primates arose, with some plesiadapiforms (e.g., carpolestids) nearer to the primate stem than others.
Geographic and Temporal Distribution
The first universally accepted primates to appear in the fossil record are the earliest members of two groups: the adapoids and the omomyoids. These groups become quite distinct over evolutionary time, filling mutually exclusive niches for the most part. However, at the start of the Eocene, the most primitive adapoids are very similar to the most primitive omomyoids.
Adapoids are called adapiforms or adapids by some and are here considered members of the superfamily Adapoidea. Omomyoids (omomyiforms or omomyids, according to some) are members of the superfamily Omomyoidea. The adapoids were mainly diurnal and herbivorous, with some achieving larger sizes than any plesiadapiforms (10 kg; 22 lbs.). By contrast, the omomyoids were mainly nocturnal, insectivorous and frugivorous, and small.
Both groups appear suddenly at the start of the Eocene, where they are present in western North America, western Europe, and India (Figure 8.6). If the primate Altiatlasius is an omomyoid, then these were also present in the Late Paleocene of North Africa. This wide dispersal of early primates was probably due to the presence of rainforest corridors extending far into northern latitudes.
In North America and Europe, both groups achieved considerable diversity in the Middle Eocene, then mostly died out at the end of that epoch. In some Eocene rock formations in the western United States, adapoids and omomyoids make up a major part of the mammalian fauna. The Eocene of India has yielded a modest diversity of euprimates, some of which are so primitive that it is difficult to know whether they are adapoids or omomyoids (or even early anthropoids).
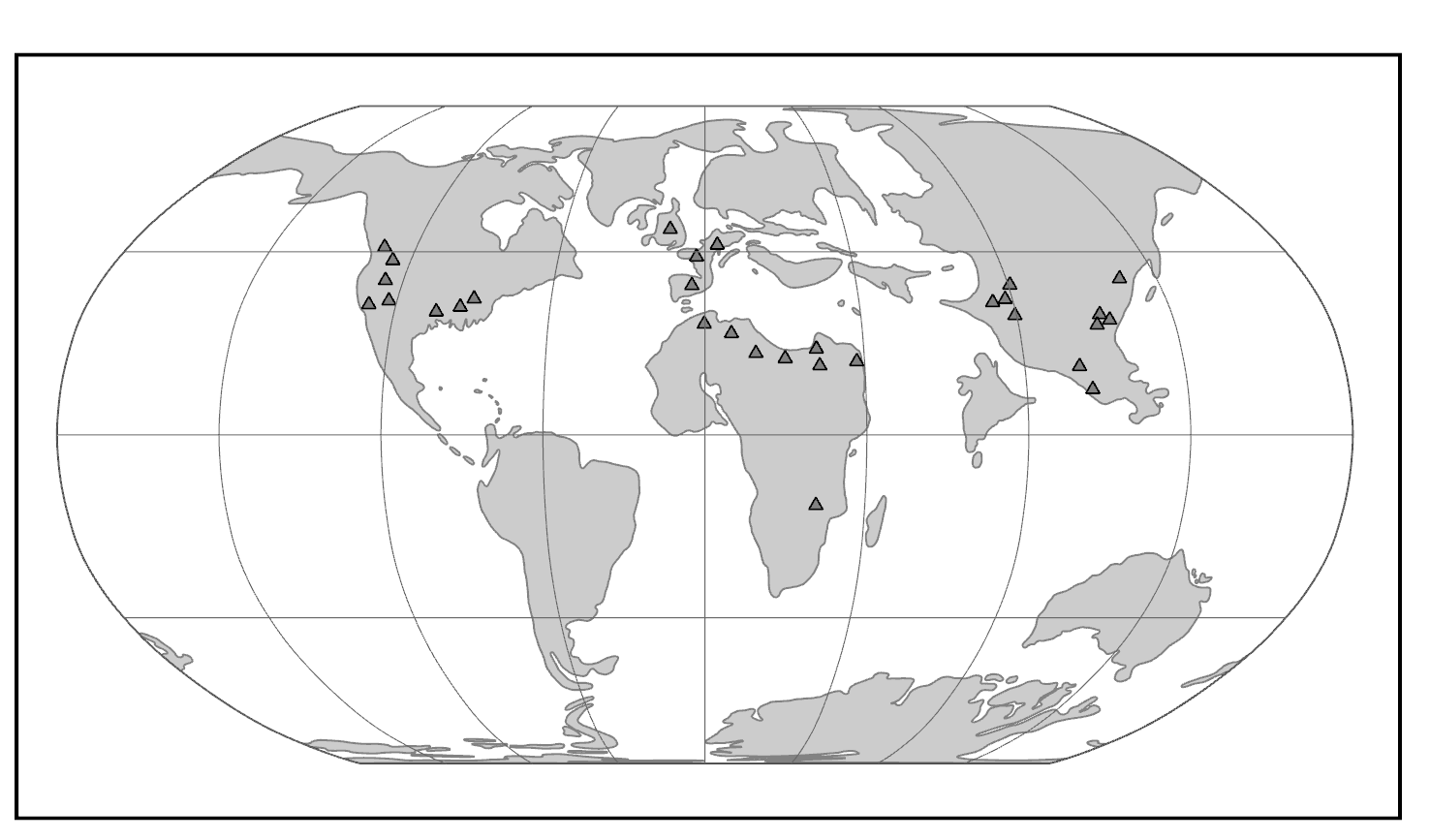 Figure \(\PageIndex{4}\): Map of the world in the Eocene, highlighting adapoid and omomyoid localities.
Figure \(\PageIndex{4}\): Map of the world in the Eocene, highlighting adapoid and omomyoid localities.Adapoids and omomyoids barely survived the Eocene-Oligocene extinctions, when colder temperatures, increased seasonality, and the retreat of rainforests to lower latitudes led to changes in mammalian biogeography. In North America, one genus originally considered an omomyoid but recently placed in the Adapoidea persisted until the Miocene: Ekgmowechashala, from various parts of North America (Rose and Rensberger 1983). This taxon has highly unusual teeth and might have been a late immigrant to North America from Asia. In Asia, one family of adapoids, the Sivaladapidae, retained considerable diversity as late as the Late Miocene. In Africa and Arabia, several fossil primates resembling extant strepsirrhines were present in the Eocene and Oligocene (approximately 34 million to 24 million years ago; Seiffert 2012).
Adapoid Diversity
Adapoids were very diverse, particularly in the Eocene of North America and Europe. They can be divided into six families, with a few species of uncertain familial relationship. As a group, adapoids have some features in common, although much of what they share is primitive. Important features include the hallmarks of euprimates: postorbital bar, flattened nails, grasping extremities, and a petrosal bulla. In addition, some adapoids retain the primitive dental formula of 2.1.4.3; that is, in each quadrant of the mouth, there are two incisors, one canine, four premolars, and three molars. In general, the incisors are small compared to the molars, but the canines are relatively large, with sexual dimorphism in some species. Their snouts are somewhat long, and evidence from cranial specimens suggests that the carotid artery branches that fed the front of the brain were variable and sometimes multiple. Cutting crests on the molars are well developed in some species, and the two halves of the mandible were fused at the midline in some species. Some adapoids were quite small (Anchomomys at a little over 100 g), and some were quite large (Magnadapis at 10 kg; 22 lbs.). Many had relatively small eyes, and in some the eyes faced somewhat laterally and/or upwardly. Furthermore, the spaces and attachment features for the chewing muscles were truly enormous in some species, suggesting that these muscles were very large and powerful. Taken together, this suggests an overall adaptive profile of diurnal herbivory. The canine sexual dimorphism in some species suggests a possible mating pattern of polygyny, as males in polygynous primate species often compete with each other for mates and have especially large canine teeth. The description that follows provides greater detail for the two best known adapoid families, the Adapidae and the Notharctidae; the additional families are summarized briefly.
The first adapoids to be described belong to the family Adapidae. This family was exclusive to Europe and includes some of the most cranially and postcranially robust primates of the Eocene (Figure 8.7). The first primate fossil ever named was Adapis, which was described by Baron Georges Cuvier between 1812 and 1822. Originally it was thought to be an ungulate and was recognized as a primate starting in the 1870s. Many adapids are known from exceptionally complete cranial and postcranial material from France and Switzerland. Unfortunately, most of the best fossils were collected in the 1800s as part of fertilizer mining operations and stratigraphic provenience of those specimens is uncertain. Furthermore, these fossils come from jumbled fissure fills, making it hard to know which bones came from the same animal.
 Figure \(\PageIndex{5}\): Representative crania of adapids (European adapoids) from the natural history museum in Montauban, France. The white scale bar is 1 cm long.
Figure \(\PageIndex{5}\): Representative crania of adapids (European adapoids) from the natural history museum in Montauban, France. The white scale bar is 1 cm long.Perhaps the best known adapoids are the Notharctidae from western North America, with a few species from Europe. Collections from Wyoming and Colorado in the United States have yielded many relatively complete skeletons—not to mention thousands of jaws with teeth—of such genera as Notharctus (Table 8.1), Cantius, and Smilodectes. The notharctids have been described as especially lemur-like (Table 8.2) on the basis of their postcranial adaptations for clinging and leaping as well as on the basis of overall cranial resemblance (Gregory 1920). The primitive Cantius has representatives in both North America and Europe.
The Cercamoniidae (e.g., Donrussellia; Table 8.1) from Europe and Asia includes some of the most primitive adapoids. The Caenopithecidae contains several genera once considered to be in Cercamoniidae, but that share an overall decrease in the length of the snout accompanied by increasing robusticity of the jaws, loss of some premolar teeth, and leaping adaptations. This family must have been very widely dispersed across the northern continents as it has representatives in the Eocene of Europe, North America, Africa, and Asia (Kirk and Williams 2011; Seiffert et al. 2009). It includes the Fayum genera Aframonius and Afradapis (Table 8.1) as well as Europolemur, Godinotia, and Darwinius (Figure 8.8) from the Messel oil shales in Germany (approximately 47.3 million years ago).
 Figure \(\PageIndex{6}\): Darwinius masillae. The slab on the left is Plate A and the slab on the right is Plate B. The parts of the skeleton in B that are outside of the dashed lines were fabricated.
Figure \(\PageIndex{6}\): Darwinius masillae. The slab on the left is Plate A and the slab on the right is Plate B. The parts of the skeleton in B that are outside of the dashed lines were fabricated.The Asiadapidae and Sivaladapidae are known from only Asia. Asiadapids are primitive adapoids from India. Postcranial elements are known from the same horizons as the described teeth, but the combination of anatomical traits in these bones makes it difficult to know if they belonged to adapoids or to omomyoids, both of which were present (Dunn et al. 2016; Rose et al. 2018). The Sivaladapidae from India, Pakistan, China, and Thailand (e.g., Sivaladapis; Table 8.1), mostly represented by jaws with teeth, persisted well into the Miocene and, in some cases, achieved a large size (4 kg; 8.82 lbs.). The sivaladapids might have evolved from the asiadapids.
Omomyoid Diversity
Like adapoids, omomyoids appeared suddenly at the start of the Eocene and then became very diverse with most species dying out before the Oligocene. Omomyoids are known from thousands of jaws with teeth, relatively complete skulls for about a half-dozen species, and very little postcranial material. Omomyoids were relatively small primates, with the largest being less than three kilograms (approximately 7 lbs.; Macrotarsiusmontanus). All known crania possess a postorbital bar, which in some has been described as “incipient closure.” Some—but not all—known crania have an elongated bony ear tube extending lateral to the location of the eardrum, a feature seen in living tarsiers and catarrhines. The anterior teeth tend to be large, with canines that are usually not much larger than the incisors. Often it is difficult to distinguish closely related species using molar morphology, but the premolars tend to be distinct from one species to another. The postcranial skeleton of most omomyoids shows hallmarks of leaping behavior reminiscent of that of tarsiers.
European omomyoids are grouped together in one family, the Microchoeridae. All microchoerids were small, had relatively large eyes, and were probably nocturnal frugivore-insectivores. Some species are known from some of the same classic fissure fill deposits as adapoids. These have yielded beautifully preserved cranial material, especially for the genus Necrolemur. The cranial morphology of Necrolemur led early paleoanthropologists to suspect they had something to do with the origin of tarsiers, a hypothesis that persists to this day (see Table 8.2). Furthermore, well-preserved ankle bones of Necrolemur and other microchoerids are strikingly like the ankle bones of tarsiers (Schmid 1979), suggesting that if there is no special relationship to tarsiers, then at least they were leaping in very similar ways. In North America, omomyoids became very diverse and abundant. In fact, omomyoids from Wyoming are sufficiently abundant and from such stratigraphically controlled conditions that they have served as strong evidence for the gradual evolution of anatomical traits over time (Rose and Bown 1984). These gradual changes have also made it difficult in some cases to establish clear boundaries between species because such boundaries usually rely on discrete differences in anatomical traits. As with European omomyoids, those in North America were probably mainly small, nocturnal, and frugivorous-insectivorous (Strait 2001). The preserved crania in some species (e.g., Shoshonius) are very like those of tarsiers (Beard and MacPhee 1994), though lacking the degree of postorbital closure. Crania in some other species (e.g., Rooneyia) are quite unusual.
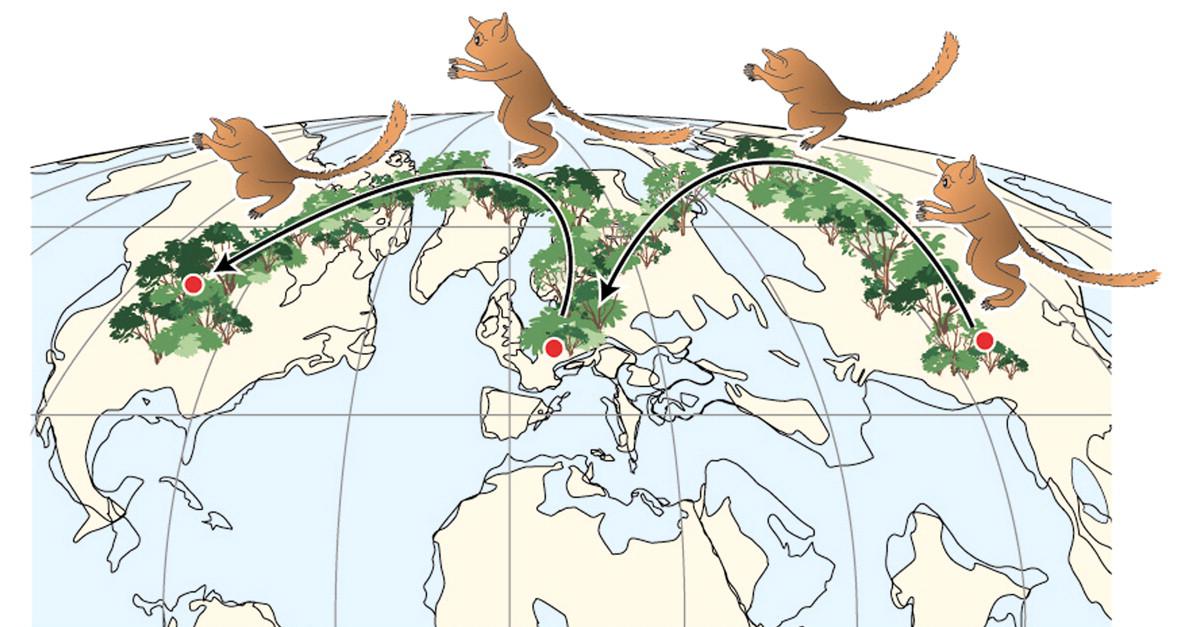 Figure \(\PageIndex{7}\): A map of the world during the earliest Eocene showing one hypothesis for the direction of dispersal of the omomyoid Teilhardina. The image also emphasizes the presence of forest corridors at high latitudes. Smith et al. 2006. Copyright (2006) National Academy of Sciences.
Figure \(\PageIndex{7}\): A map of the world during the earliest Eocene showing one hypothesis for the direction of dispersal of the omomyoid Teilhardina. The image also emphasizes the presence of forest corridors at high latitudes. Smith et al. 2006. Copyright (2006) National Academy of Sciences.North American omomyoids are grouped into one family, Omomyidae, with two subfamilies: Anaptomorphinae and Omomyinae. The anaptomorphines were mostly smaller and more generalized whereas some omomyines achieved larger sizes (1–2 kg; 2–4 lbs.) and some were more specialized.
Teilhardina (Figure 8.9; Table 8.1) is one of the earliest and arguably the most primitive of omomyoids. Teilhardina has several species, most of which are from North America, with one from Europe (T. belgica) and one from Asia (T. asiatica). The species of this genus are anatomically similar and the deposits from which they are derived are roughly contemporaneous. Thus, this small primate likely dispersed across the northern continents very rapidly (Smith et al. 2006).