
8.1.4: The Emergence of Modern Primate Groups
- Last updated
- Save as PDF
- Page ID
- 136422
Origins of Crown Strepsirrhines
Until the turn of this century, very little was known about the origins of the crown (living) strepsirrhines. The Quaternary record of Madagascar contains many amazing forms of lemurs, including giant sloth-like lemurs, lemurs with perhaps monkey-like habits, lemurs with koala-like habits, and even a giant aye-aye (Godfrey and Jungers 2002). However, in Madagascar early Tertiary continental sediments are lacking, and there is no record of lemur fossils before the Pleistocene.
The fossil record of galagos is slightly more informative. Namely, there are Miocene African fossils that are very likely progenitors of lorisids (Simpson 1967). However, these are much like modern galagos and do not reveal anything about the relationship between crown strepsirrhines and Eocene fossil primates (but see below regarding Propotto). A similar situation exists for lorises in Asia: there are Miocene representatives, but these are substantially like modern lorises.
In 2003, the description of two new fossil genera from the Birket Qarun locality in the Fayum Basin of Egypt provided the “smoking gun” for the origin of crown strepsirrhines (Seiffert et al. 2003). Saharagalago and Karanisia are two small primates that bear clear affinities with crown strepsirrhines to the exclusion of other primates. Most striking is a lower canine of Karanisia that clearly sat within a strepsirrhine-style toothcomb. Recently, several other African primates have been recognized as having strepsirrhine affinities (Seiffert 2012). These include Azibius and Algeripithecus from Algeria, Djebelemur from Tunisia, Omanodon and Shizarodon from Oman, and Namaia from Namibia (see Marivaux et al. 2013). These important fossil primates are mainly known from teeth and jaws. The enigmatic Fayum primate Plesiopithecus is known from a single skull that has been compared to aye-ayes and to lorises (Godinot 2006; Simons and Rasmussen 1994a).
The now-recognized diversity of stem strepsirrhines from the Eocene and Oligocene of Afro-Arabia is strong evidence to suggest that strepsirrhines originated in that geographic area. This implies that lorises dispersed to Asia subsequent to an African origin. It is unknown what the first strepsirrhines in Madagascar were like. However, it seems likely that the lemuriform-lorisiform split occurred in continental Africa, followed by dispersal of lemuriform stock to Madagascar. Recent evidence suggests that Propotto, a Miocene primate from Kenya originally described as a potto antecedent, actually forms a clade with Plesiopithecus and the aye-aye; this might suggest that strepsirrhines dispersed to Madagascar from continental Africa more than once (Gunnell et al. 2018).
The Fossil Record of Tarsiers
Tarsiers are so unusual that they fuel major debates about primate taxonomy. Tarsiers today are moderately diverse but geographically limited and not very different in their ecological habits—especially considering that the split between them and their nearest living relative probably occurred over 50 million years ago. If omomyoids are excluded, then the fossil record of tarsiers is very limited. Two fossil species from the Miocene of Thailand have been placed in the genus Tarsius, as has an Eocene fossil from China (Beard et al. 1994). These, and Xanthorhysis from the Eocene of China, are all very tarsier-like. In fact, it is striking that Tarsius eocaenus from China was already so tarsier-like as early as the Eocene. This suggests that tarsiers achieved their current morphology very early on in their evolution and have remained more or less the same while other primates changed dramatically. Two additional genera, Afrotarsius from the Oligocene of Egypt and Libya and Afrasia from the Eocene of Myanmar, have also been implicated in tarsier origins, though the relationship between them and tarsiers is unclear (Chaimanee et al. 2012). More recently, a partial skeleton of a small Eocene primate from China, Archicebus achilles (dated to approximately 55.8 million to 54.8 million years ago), was described as the most basal tarsiiform (Ni et al. 2013). This primate is reconstructed as a diurnal insectivore and an arboreal quadruped that did some leaping—but not to the specialized degree seen in living tarsiers. The anatomy of the eye in living tarsiers suggests that their lineage passed through a diurnal stage, so Archicebus (and diurnal omomyoids) might represent such a stage.
Climate Change and the Paleogeography of Modern Primate Origins
 Figure \(\PageIndex{1}\): Map of key localities of early anthropoids. Note that the landmasses are in their current configuration.
Figure \(\PageIndex{1}\): Map of key localities of early anthropoids. Note that the landmasses are in their current configuration.Changing global climate has had profound effects on primate dispersal patterns and ecological habits over evolutionary time. Primates today are strongly tied to patches of trees and particular plant parts such as fruits, seeds, and immature leaves. It is no surprise, then, that the distribution of primates mirrors the distribution of forests. Today, primates are most diverse in the tropics and especially in tropical rainforests. Global temperature trends across the Tertiary have affected primate ranges. Following the Cretaceous-Tertiary extinction event, cooler temperatures and greater seasonality characterized the Paleocene. In the Eocene, temperatures (and probably rainfall) increased globally and rainforests likely extended to very high latitudes. During this time, euprimates became very diverse. With cooling and increased aridity at the end of the Eocene, many primate extinctions occurred in the northern continents and the surviving primates were confined to lower latitudes in South America, Afro-Arabia, Asia, and southern Europe. Among these survivors are the progenitors of the living groups of primates: lemurs and lorises, tarsiers, New World monkeys, Old World monkeys, and apes (Figure 8.10).
Competing Hypotheses for the Origin of Anthropoids
There is considerable debate among paleoanthropologists as to the geographic origins of anthropoids. In addition, there is debate regarding the source group for anthropoids. Three different hypotheses have been articulated in the literature. These are the adapoid origin hypothesis, the omomyoid origin hypothesis, and the tarsier origin hypothesis (Figure \(\PageIndex{2}\)).
 Figure \(\PageIndex{2}\): Competing trees for anthropoid origins. Branch lengths are not to scale. Note that the position of strepsirrhines is not necessarily specified by the omomyoid origin and tarsier origin models.
Figure \(\PageIndex{2}\): Competing trees for anthropoid origins. Branch lengths are not to scale. Note that the position of strepsirrhines is not necessarily specified by the omomyoid origin and tarsier origin models.Adapoid Origin Hypothesis
Resemblances between some adapoids and some extant anthropoids include fusion of the mandibular symphysis, overall robusticity of the chewing system, overall large body size, features that signal a diurnal lifestyle (like relatively small eye sockets), and details of ankle bone morphology. Another feature in common is canine sexual dimorphism, which is present in some species of adapoids (probably) and in several species of anthropoids.
These features led some paleoanthropologists in the last half of the 20th century to suggest that anthropoids arose from adapoid stock (Gingerich 1980; Simons and Rasmussen 1994b). One of the earliest supporters of the link between adapoids and anthropoids was Hans Georg Stehlin, who described much of the best material of adapoids and who compared these Eocene primates to South American monkeys (Stehlin 1912). In more recent times, the adapoid origin hypothesis was reinforced by resemblances between these European adapoids (especially Adapis and Leptadapis) and some early anthropoids from the Fayum Basin (e.g., Aegyptopithecus, see below; Table 8.1).
Unfortunately for the adapoid hypothesis, most of the shared features listed above probably arose independently in the two groups as adaptations to a diet of hard and/or tough foods. For example, fusion of the mandibular symphysis likely evolved as a means to strengthen the jaw against forces that would pull the two halves away from each other, in the context of active chewing muscles on both sides of the head generating great bite forces. This context would also favor the development of robust jaws, large chewing muscles, shorter faces, and some other features shared by some adapoids and some anthropoids.
As older and more primitive anthropoids were found in the Fayum Basin, it became clear that the earliest anthropoids from Africa do not possess these features of jaw robusticity (Seiffert et al. 2009). Furthermore, many adapoids never evolved these features. Fusion of the mandibular symphysis in adapoids is actually quite different from that in anthropoids and probably occurred during juvenile development in the former (Beecher 1983; Ravosa 1996). Eventually, the adapoid origin hypothesis fell out of favor among most paleoanthropologists, although the description of Darwinius is a recent revival of that idea (Franzen et al. 2009; but see Seiffert et al. 2009, Williams et al. 2010b).
Omomyoid Origin Hypothesis
Similarities in cranial and hindlimb morphology between some omomyoids and extant tarsiers have led to the suggestion that tarsiers arose from some kind of omomyoid. In particular, Necrolemur has many features in common with tarsiers, as does the North American Shoshonius, which is known from a few beautifully preserved (although distorted) crania. Tarsiers and Shoshonius share exclusively some features of the base of the cranium; however, Shoshonius does not have any sign of postorbital closure and it lacks the bony ear tube of tarsiers. Nevertheless, some of the resemblances between some omomyoids and tarsiers suggest that tarsiers might have originated from within the Omomyoidea (Beard 2002; Beard and MacPhee 1994). In this scenario, although living tarsiers and living anthropoids might be sister taxa, they might have evolved from different omomyoids, possibly separated from each other by more than 50 million years of evolution, or anthropoids evolved from some non-omomyoid fossil group. The arguments against the omomyoid origin hypothesis are essentially the arguments for the tarsier origin hypothesis (see below). Namely, tarsiers and anthropoids share many features (especially of the soft tissues) that must have been retained for many millions of years or must have evolved convergently in the two groups. Furthermore, a key hard-tissue feature shared between the two extant groups, the postorbital septum, was not present in any omomyoid. Therefore, that feature must have arisen convergently in the two extant groups or must have been lost in omomyoids. Neither scenario is very appealing, although recent arguments for convergent evolution of the postorbital septum in tarsiers and anthropoids have arisen from embryology and histology of the structure (DeLeon et al. 2016).
Tarsier Origin Hypothesis
Several paleoanthropologists have suggested that there is a relationship between tarsiers and anthropoids to the exclusion of omomyoids and adapoids (e.g., Cartmill and Kay 1978; Ross 2000; Williams and Kay 1995). As mentioned, tarsiers and anthropoids today share several traits. These include many soft-tissue features related to the olfactory system such as the loss of a hairless external nose and loss of the median cleft running from the nose to the mouth (possessed by strepsirrhines). Also included are aspects of the visual system such as the loss of a reflective layer at the back of the eye, similarities in carotid circulation to the brain, and mode of placentation. Unfortunately, none of these can be assessed directly in fossils. Some bony similarities between tarsiers and anthropoids include an extra air-filled chamber below the middle ear cavity, reduced bones within the nasal cavity, and substantial postorbital closure; these can be assessed in fossils, but the distribution of these traits in omomyoids does not yield clear answers. Furthermore, several of the similarities between tarsiers and anthropoids are probably due to similarities in the sensory systems, which might have evolved in parallel for ecological reasons. Although early attempts to resolve the crown primates with molecular data were sometimes equivocal or in disagreement with one another, more recent analyses (including those of short interspersed elements) suggest that tarsiers and anthropoids are sister groups to the exclusion of lemurs and lorises (Williams et al. 2010a). However, this does not address omomyoids, all of which are far too ancient for DNA extraction.
The above three hypotheses are not the only possibilities for anthropoid origins. It may be that anthropoids are neither the closest sister group of tarsiers, nor evolved from adapoids or omomyoids. In recent years, two new groups of Eocene Asian primates have been implicated in the origin of anthropoids: the eosimiids and the amphipithecids. It is possible that one or the other of these two groups gave rise to anthropoids. Regardless of the true configuration of the tree for crown primates, the three major extant groups probably diverged from each other quite long ago (Seiffert et al. 2004).
Early Anthropoid Fossils in Africa
 Figure \(\PageIndex{3}\): Egyptian workers sweeping Quarry I in the Fayum Basin (2004). This is a technique called wind harvesting that removes the desert crust and permits wind to blow out fine sediment and reveal fossils.
Figure \(\PageIndex{3}\): Egyptian workers sweeping Quarry I in the Fayum Basin (2004). This is a technique called wind harvesting that removes the desert crust and permits wind to blow out fine sediment and reveal fossils. Figure \(\PageIndex{4}\): Elwyn Laverne Simons excavating Aegyptopithecus in the Fayum Basin.
Figure \(\PageIndex{4}\): Elwyn Laverne Simons excavating Aegyptopithecus in the Fayum Basin.The classic localities yielding the greatest wealth of early anthropoid fossils are those from the Fayum Basin in Egypt (Simons 2008; Figure 8.12). The Fayum is a veritable oasis of fossil primates in an otherwise rather spotty early Tertiary African record. Since the 1960s, teams led by E. L. Simons have discovered several new species of early anthropoids, some of which are known from many parts of the skeleton and several individuals (Figure 8.13).
The Fayum Jebel Qatrani Formation and Birket Qarun Formation between them have yielded a remarkable array of terrestrial, arboreal, and aquatic mammals. These include ungulates, bats, sea cows, elephants, hyraces, rodents, whales, and primates. Also, many other vertebrates, like water birds, were present. The area at the time of deposition (Late Eocene through Early Oligocene) was probably very wet, with slow-moving rivers, standing water, swampy conditions, and lots of trees (see Bown and Kraus 1988). In short, it was an excellent place for primates.
General Morphology of Anthropoids
The anthropoids known from the Fayum (and their close relatives from elsewhere in East Africa and Afro-Arabia) bear many of the anatomical hallmarks of extant anthropoids; however, there are primitive forms in several families that lack one or more anthropoid traits. All Fayum anthropoids known from skulls possess postorbital closure, most had fused mandibular symphyses, and most had ring-like ectotympanic bones. Tooth formulae were generally either 2.1.3.3 or 2.1.2.3. Fayum anthropoids ranged in size from the very small Qatrania and Biretia (less than 500 g) to the much-larger Aegyptopithecus (approximately 7 kg; 15 lbs.). Fruit was probably the main component of the diet for most or all of the anthropoids, with some of them supplementing with leaves (Kay and Simons 1980; Teaford et al. 1996; Kirk and Simons 2001). Most Fayum anthropoids were probably diurnal above-branch quadrupeds. Some of them (e.g., Apidium; Table 8.1) were probably very good leapers (Gebo and Simons 1987), but none show specializations for gibbon-style suspensory locomotion. Some of the Fayum anthropoids are known from hundreds of individuals, permitting the assessment of individual variation, sexual dimorphism, and in some cases growth and development. The description that follows provides greater detail for the two best known Fayum anthropoid families, the Propliopithecidae and the Parapithecidae; the additional families are summarized briefly.
Fayum Anthropoid Families
The Propliopithecidae (e.g., Pliopithecus; Table 8.1) include the largest anthropoids from the fauna, and they are known from several crania and some postcranial elements. They have been suggested to be stem catarrhines (Old World Monkeys and apes), although perhaps near the split between catarrhines and platyrrhines. The best known propliopithecid is Aegyptopithecus, known from many teeth, crania, and postcranial elements. The crania (Figure 8.14) show substantial morphological variation between individuals, some of which might be due to sexual dimorphism and a polygynous mating system (Simons et al. 2007). Aegyptopithecus was large (greater than 7 kg; 15 lbs.) with prominent attachments for the chewing muscles and with low, rounded, and well-buttressed molars. The snout is long and the canines are large. There is only partial development of a bony ear tube (Simons et al. 2007). The known long bones of Aegyptopithecus are quite robust, and the skeleton suggests that this animal was a generalized arboreal quadruped with no strong specialization for suspension (Gebo and Simons 1987).
 Figure \(\PageIndex{5}\): Female and male skull material for Aegyptopithecus zeuxis. The female is on the left. The mandibles are not associated with the crania. Simons et al. 2007. Copyright (2007) National Academy of Sciences.
Figure \(\PageIndex{5}\): Female and male skull material for Aegyptopithecus zeuxis. The female is on the left. The mandibles are not associated with the crania. Simons et al. 2007. Copyright (2007) National Academy of Sciences.An extremely abundant and unusual family of anthropoids from the Fayum is the Parapithecidae. Parapithecids have the primitive anthropoid tooth formula of 2.1.3.3 and they have very large conules on the upper molars and premolars. The parapithecid Apidium is known from many jaws with teeth, crushed and distorted crania, and several skeletal elements. The molars of Apidium had low, rounded cusps and thick enamel, suggesting a frugivorous diet. Apidium was probably a good leaper, having tightly apposed distal tibia and fibula as well as a narrow distal femur with a deep patellar groove (Fleagle and Simons 1995). Parapithecus is known from cranial material including a beautiful, undistorted cranium. This genus shows extreme reduction of the incisors, including complete absence of the lower incisors in P. grangeri (Simons 2001). This trait is unique among primates. Biretia is a primitive member of the family that has the primitive trait of an unfused mandibular symphysis. Parapithecids were once thought to be the ancestral stock of platyrrhines; however, their platyrrhine-like features are probably just primitive retentions and the most conservative approach is to consider them stem anthropoids.
The Oligopithecidae share the catarrhine tooth formula of 2.1.2.3 as well as having a canine honing complex that involves the anterior lower premolar. The postcranial elements known for the group suggest generalized arboreal quadrupedalism. The best known member, Catopithecus, is known from crania that demonstrate a postorbital septum and from mandibles that lack symphyseal fusion (Simons and Rasmussen 1996). The jaws are deep, with broad muscle attachment areas and crested teeth. Catopithecus was probably a little less than a kilogram in weight.
The Proteopithecidae had an overall primitive dentition that includes three premolars per quadrant and a generalized skeleton; they are considered stem anthropoids. The best known genus, Proteopithecus, is represented by dentitions, crania, and postcranial elements that suggest a diet of mostly fruit and a generalized style of locomotion, including arboreal quadrupedalism with some leaping (Simons and Seiffert 1999). It weighed about a kilogram.
Other genera of putative anthropoids from the Fayum include the very poorly known Arsinoea, the contentious Afrotarsius, and the enigmatic Nosmips. The last of these possesses traits of several major primate clades and defies classification (Seiffert et al. 2010).
Early Anthropoid Fossils in Asia
For the last half of the 1900s, researchers believed that Africa was the unquestioned homeland of early anthropoids (see Fleagle and Kay 1994). However, two very different groups of primates from Asia soon began to change that. One was an entirely new discovery (Eosimiidae), and the other was a poorly known group discovered decades prior (Amphipithecidae). Soon, attention on anthropoid origins began to shift eastward (see Ross and Kay 2004, Simons 2004). If anthropoids arose in Asia instead of Africa, then this implies that the African early anthropoids either emigrated from Asia or evolved their anthropoid traits in parallel with living anthropoids.
Eosimiids
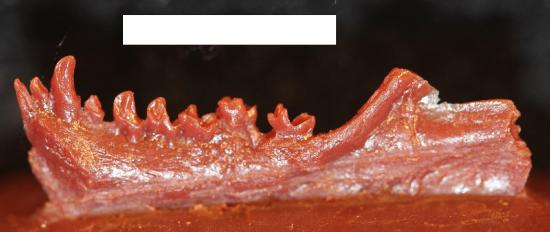 Figure \(\PageIndex{6}\): Cast of the right half of the mandible of Eosimias centennicus, type specimen. The white scale bar is 1 cm long.
Figure \(\PageIndex{6}\): Cast of the right half of the mandible of Eosimias centennicus, type specimen. The white scale bar is 1 cm long.First described in the 1990s, the eosimiids are best represented by Eosimias (Table 8.1). This “dawn monkey” is known from relatively complete jaws with teeth, a few small fragments of the face, and some postcranial elements (Beard et al. 1994; Beard et al. 1996; Gebo et al. 2000). The lower jaw is distinctive in being very deep relative to its length and breadth, as in some early Fayum anthropoids (Figure 8.15). It also has pointed incisors that are about the same size as the canines. The lower premolars are crowded together and they are set obliquely in the jaw. This last trait, also present in another eosimiid called Bahinia, was described as a shared derived trait with anthropoids, though it is also present in some adapoids (e.g., Afradapis from North Africa and Asiadapis [Table 8.1] from India). The mandibular symphysis is vertically inclined, as in some anthropoids, but is unfused. Overall, Eosimias was small, no heavier than a can of soda pop. Eosimias (along with the other less-well-known genera in its family) bears some resemblance to tarsiers as well as anthropoids. The shared features with anthropoids are mainly jaw shape and details of dental morphology. Unfortunately, no good crania are known for this family and the anatomy of, for example, the posterior orbital margin could be very revealing as to higher-level relationships.
Amphipithecids
Amphipithecids are small- to medium-size primates (up to 10 kg; 22 lbs.). Most are from the Eocene Pondaung Formation in Myanmar (Early–Middle Eocene), but one genus is known from Thailand. They were first discovered in the 1910s, and all of the specimens discovered in the first half of the 20th century were fragmentary jaws with teeth that were mostly worn down. Nevertheless, some dental similarities with anthropoids were noted early on. These include deep jaws and wide basins that separate low molar cusps. Starting in the 1970s, intensive collecting efforts in Myanmar yielded new material for the best known genera Pondaungia and Amphipithecus (Ciochon and Gunnell 2002; Table 8.1). Soon, another genus was discovered: Myanmarpithecus. It bears some resemblance to the other genera but has longer molar crests, suggesting a higher degree of folivory (Kay et al. 2004). Another amphipithecid, Siamopithecus from Thailand, has very rounded molars and was probably a seed-eater (Figure 8.16). In addition to teeth and jaws, some cranial fragments, ankle material, and ends of postcranial bones have been found for Pondaungia. There are important resemblances between the postcranial bones of Pondaungia and those of adapoids, suggesting adapoid affinities for the amphipithecidae. This would imply that the resemblances with anthropoids in the teeth are convergent, based on similarities in diet (see Ciochon and Gunnell 2002). Unfortunately, the association between postcranial bones and teeth is not definite. With other primates in these faunas (including eosimiids), one cannot be certain that the postcranial bones belong with the teeth. Perhaps, as suggested by some, some of the bones belong to a sivaladapid (or asiadapid) and others belong to an early anthropoid (Beard et al. 2007; Marivaux et al. 2003). Additional well-associated material of amphipithecids would help to clear up this uncertainty.
 Figure \(\PageIndex{7}\): Casts of representative amphipithecid material. A, Pondaungia cotteri right lower jaw fragment with m2 and m3. B, Siamopithecus eocaenus right upper jaw fragment with p4-m3. C, S. eocaenus right lower jaw fragment with partial m1, m2, and m3 in lateral view; note the great depth of this jaw. D, same as in C, but occlusal view. White scale bars are 1 cm long; the scale is the same for A, B, and D.
Figure \(\PageIndex{7}\): Casts of representative amphipithecid material. A, Pondaungia cotteri right lower jaw fragment with m2 and m3. B, Siamopithecus eocaenus right upper jaw fragment with p4-m3. C, S. eocaenus right lower jaw fragment with partial m1, m2, and m3 in lateral view; note the great depth of this jaw. D, same as in C, but occlusal view. White scale bars are 1 cm long; the scale is the same for A, B, and D.Platyrrhine Dispersal to South America
Today there is an impressive diversity of primates in South and Central America. These are considered to be part of a single clade, the Platyrrhini. Primates colonized South America sometime in the Eocene from an African source. In the first half of the 20th century, the source of platyrrhines was a matter of major debate among paleontologists, with some favoring a North American origin (e.g., Simpson 1940).
Part of the reason for this debate is that South America was an island in the Eocene. Primates needed to cross open ocean to get there from either North America or Africa, although the distance from the former was shorter. Morphology yields clues to platyrrhine origins. The first known primates in South America have more in common morphologically with African primates than with North American ones. At the time, anthropoids were popping up in North Africa, whereas the only euprimates in North America were adapoids and omomyoids. Despite lacking a bony ear tube, early platyrrhines shared a great deal with other anthropoids, including full postorbital closure and fusion of the mandibular symphysis.
The means by which a population of small North African primates managed to disperse across the Atlantic and survive to colonize South America remains a mystery. The most plausible scenario is one of rafting. That is, primates must have been trapped on vegetation that was blown out to sea by a storm. The vegetation then became a sort of life raft, which eventually landed ashore, dumping its passengers in South America. Rodents probably arrived in South America in the same way (Antoine et al. 2012).
Once ashore, platyrrhines must have crossed South America fairly rapidly because the earliest-known primates from that continent are from Peru (Bond et al. 2015). Soon after that, platyrrhines were in Bolivia, namely Branisella. By the Miocene, platyrrhines were living in extreme southern Argentina and were exploiting a variety of feeding niches. The Early Miocene platyrrhines were all somewhat primitive in their morphology, but some features that likely arose by ecological convergence suggest (to some) relationships with extant platyrrhine families. This has led to a lively debate about the pattern of primate evolution in South America (Kay 2015; Kay and Fleagle 2010; Rosenberger 2010). By the Middle Miocene, clear representatives of modern families were present in a diverse fauna from La Venta, Colombia (Wheeler 2010). The Plio-Pleistocene saw the emergence of giant platyrrhines as well as several taxa of platyrrhines living on Caribbean islands (Cooke et al. 2016).
The story of platyrrhines seems to be one of amazing sweepstakes dispersal, followed by rapid diversification and widespread geographic colonization of much of South America. After that, dramatic extinctions resulted in the current, much smaller geographic distribution of platyrrhines. These extinctions were probably caused by changing climates, leading to the contraction of forests. Platyrrhines dispersed to the Caribbean and to Central America, with subsequent extinctions in those regions that might have been related to interactions with humans. Unlike anthropoids of the Old World, platyrrhines do not seem to have evolved any primarily terrestrial forms and so have always been highly dependent on forests.
SPECIAL TOPIC: JONATHAN PERRY AND PRIMATES OF THE EXTREME SOUTH
Many primates are very vulnerable to ecological disturbance because they are heavily dependent on fruit to eat and trees to live in. This is one reason why so many primates are endangered today and why many of them went extinct due to climatic and vegetational changes in the past. Jonathan Perry’s paleontological research focuses on primates that lived on the edge of their geographic distribution. This research has taken him to two extremes in the Americas: extreme southern Patagonia and the Canadian prairies.
Santa Cruz Province in Argentina is as far south as primates have ever lived. The Santa Cruz fauna of the Miocene has yielded a moderate diversity of platyrrhines, each with slightly different dietary adaptations. These include Homunculus (Table 8.1), first described by Florentino Ameghino in 1891 (Figure 8.17). Recent fieldwork by Perry and colleagues in Argentina has revealed several skulls of Homunculus as well as many parts of the skeleton (Kay et al. 2012). The emerging profile of this extinct primate is one of a dedicated arboreal quadruped that fed on fruits and leaves. Many of the foods eaten by Homunculus must have been very tough and were probably covered and impregnated with grit; we suspect this because the cheek teeth are very worn down, even in young individuals, and because the molar tooth roots were very large, presumably to resist strong bite forces (Perry et al. 2010, 2014).
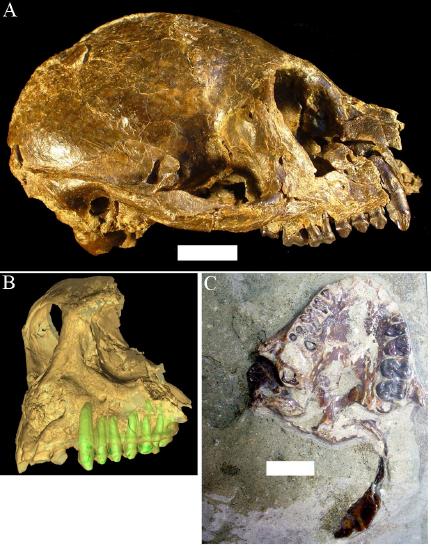 Figure \(\PageIndex{1}\): Representative specimens of Homunculus patagonicus. A, adult cranium in lateral view. B, adult cranium surface reconstructed from microCT scans, with the teeth segmented out. C, juvenile cranium. White scale bars are 1cm long.
Figure \(\PageIndex{1}\): Representative specimens of Homunculus patagonicus. A, adult cranium in lateral view. B, adult cranium surface reconstructed from microCT scans, with the teeth segmented out. C, juvenile cranium. White scale bars are 1cm long.Perry began working in Argentina while a graduate student at Duke University. He participated as a field assistant in a team led by his Ph.D. advisor, Richard F. Kay, and Argentine colleagues Sergio F. Vizcaíno and M. Susana Bargo. Most of the localities examined belong to a suite of beach sites known since the 1800s and visited by many field parties from various museums in the early 1900s. Since 2003, their international team of paleontologists from the U.S. and Argentina has visited these localities every single year (Figure 8.18). Over time, new fossils and new students have led to new projects and new approaches, including the use of microcomputed tomography (microCT) to visualize and analyze internal structures of the skeleton.
 Figure \(\PageIndex{2}\): Field localities in Argentina and Canada. A, Cañadon Palos locality, coastal Santa Cruz Province, Argentina. B, Swift Current Creek locality, southwest Saskatchewan, Canada.
Figure \(\PageIndex{2}\): Field localities in Argentina and Canada. A, Cañadon Palos locality, coastal Santa Cruz Province, Argentina. B, Swift Current Creek locality, southwest Saskatchewan, Canada.