
10.3: Homo Erectus
- Page ID
- 177681
BIOLOGICAL AND CULTURAL INNOVATIONS
Homo erectus Anatomy
A ridge on the occipital bone in the back of the skull.
A thickened area along the top of the skull.
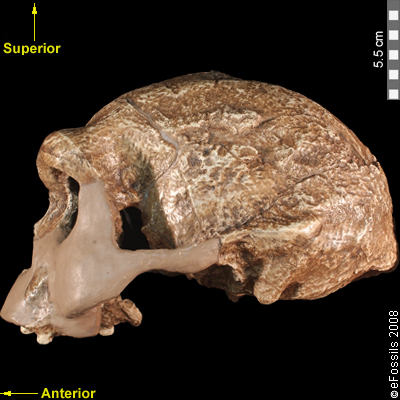 Figure \(\PageIndex{1}\): Replica of Homo erectus from Java, Indonesia. This cranium (known as Sangiran 17) dates to approximately 1.3 million to 1 million years ago. Note the large brow ridges and the occipital torus that gives the back of the skull a squared-off appearance.
Figure \(\PageIndex{1}\): Replica of Homo erectus from Java, Indonesia. This cranium (known as Sangiran 17) dates to approximately 1.3 million to 1 million years ago. Note the large brow ridges and the occipital torus that gives the back of the skull a squared-off appearance.Homo erectus in Africa
Regional Discoveries Outside Africa
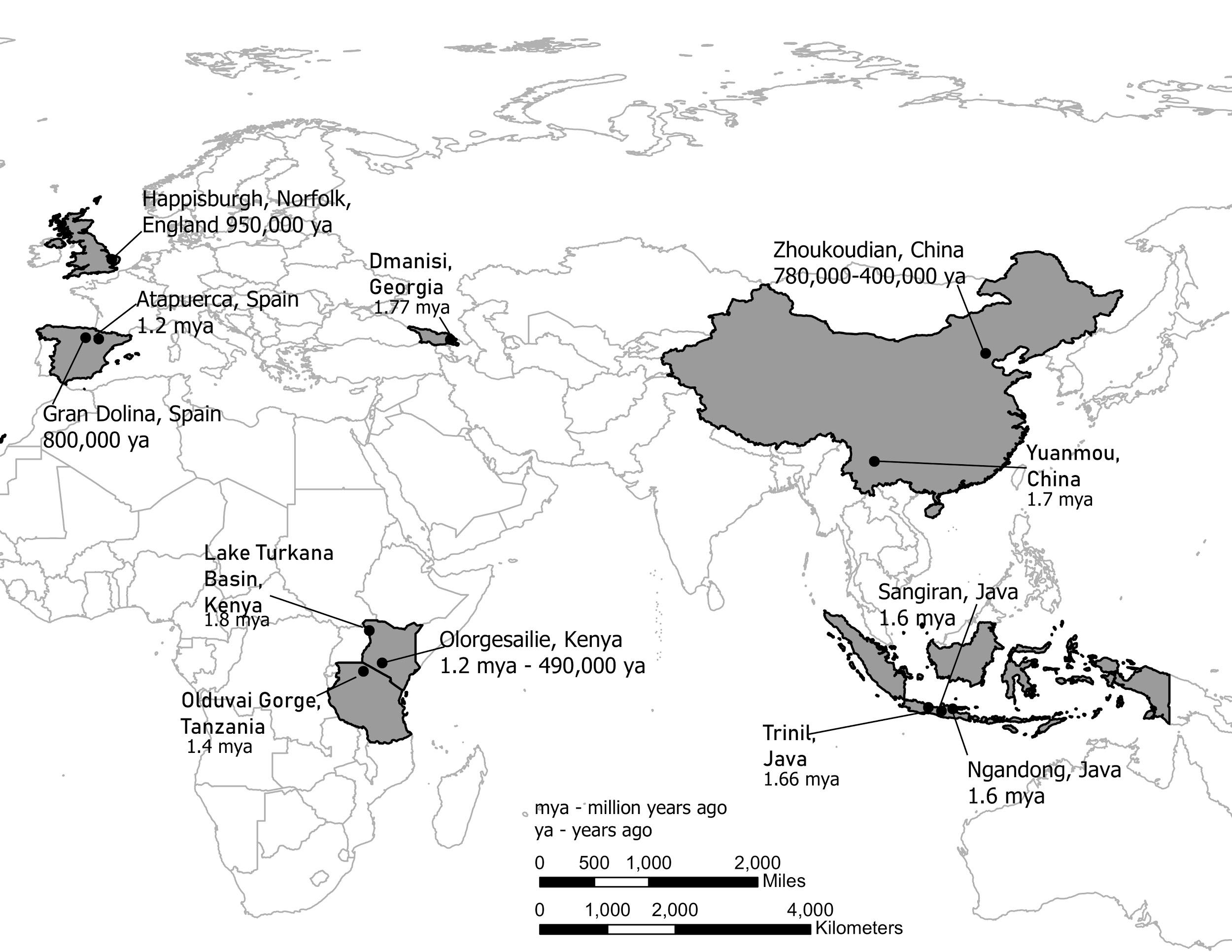 Figure \(\PageIndex{2}\): Map showing the locations of Homo erectus fossils around Africa and Eurasia.
Figure \(\PageIndex{2}\): Map showing the locations of Homo erectus fossils around Africa and Eurasia.Indonesia
China
Western Eurasia
Europe
LIFEWAYS
Now, our examination of Homo erectus will turn to its lifeways—how the species utilized its environment in order to survive. This includes making inferences about diet, technology, life history, environments occupied, and perhaps even social organization. As will be apparent, Homo erectus shows significant cultural innovations in these areas, some that you will probably recognize as more “human-like” than any of the hominins previously covered.
Tool Technology: Acheulean Tool Industry
In early African sites associated with Homo erectus, stone tools such as flakes and choppers identified to the Oldowan Industry dominate. Starting at about 1.5 million years ago, some Homo erectus populations began making different forms of tools. These tools–classified together as constituting the Acheulean tool industry–are more complex in form and more consistent in their manufacture. Unlike the Oldowan tools, which were cobbles modified by striking off a few flakes, Acheulean toolmakers carefully shaped both sides of the tool. This type of technique, known as bifacial flaking, requires more planning and skill on the part of the toolmaker; he or she would need to be aware of principles of symmetry when crafting the tool. One of the most common tool forms, the handaxe, is shown in Figure \(\PageIndex{3}\). As with the tool illustrated below, handaxes tend to be thicker at the base and then come to a rounded point at the tip. Besides handaxes, forms such as scrapers, cleavers, and flake tools are present at Homo erectus sites.
Tool industry characterized by teardrop-shaped stone handaxes flaked on both sides.
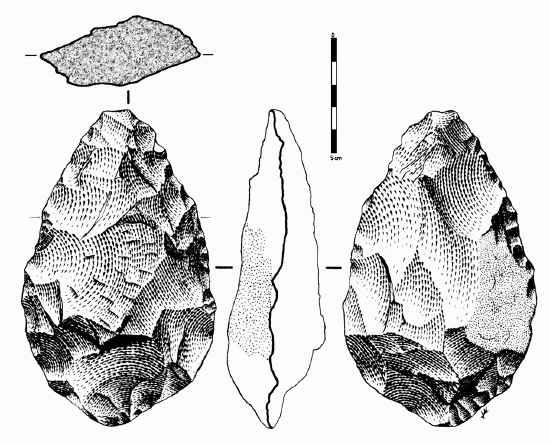 Figure \(\PageIndex{3}\): Drawing of an Acheulean handaxe. This specimen is from Spain. When drawing a stone tool, artists typically show front and back faces, as well as top and side profiles.
Figure \(\PageIndex{3}\): Drawing of an Acheulean handaxe. This specimen is from Spain. When drawing a stone tool, artists typically show front and back faces, as well as top and side profiles.One striking aspect of Acheulean tools is their uniformity. They are more standardized in form and mode of manufacture than the earlier Oldowan tools. For example, the aforementioned handaxes vary in size, but they are remarkably consistent in regard to their shape and proportions. They were also an incredibly stable tool form over time—lasting well over a million years with little change.
Curiously, the Acheulean tools so prominent at African sites are mostly absent in Homo erectus sites in East Asia. Instead, Oldowan-type choppers and scrapers are found at those sites. If this technology seemed to be so important to African Homo erectus, why didn’t East Asian Homo erectus also use the tools? One reason could be environmental differences between the two regions. Perhaps the rocks available in Asia weren’t of the material suitable for making the Acheulean handaxes. It has been suggested that Asian Homo erectus populations used perishable material such as bamboo to make tools. Another possibility is that Homo erectus (or even an earlier hominin) migrated to East Asia before the Acheulean technology developed in Africa. The recent discovery of the 2.1 million-year-old tools in China gives credence to this last explanation.
Tool Use and Cognitive Abilities of Homo erectus
What (if anything) do the Acheulean tools tell us about the mind of Homo erectus? Clearly, they took a fair amount of skill to manufacture. Apart from the actual shaping of the tool, other decisions made by toolmakers can reveal their use of foresight and planning. Did they just pick the most convenient rocks to make their tools, or did they search out a particular raw material that would be ideal for a particular tool? Analysis of Acheulean stone tools suggest that at some sites, the toolmakers selected their raw materials carefully—traveling to particular rock outcrops to quarry stones and perhaps even removing large slabs of rock at the quarries to get at the most desirable material. Such complex activities would require advanced planning. They also likely required cooperation and communication with other individuals, as such actions would be difficult to carry out solo. However, other Homo erectus sites lack evidence of such selectivity; instead of traveling even a short distance for better raw material, the hominins tended to use what was available in their immediate area (Shipton et al. 2018).
In contrast to Homo erectus tools, the tools of early modern Homo sapiens during the Upper Paleolithic display tremendous diversity across regions and time periods. Additionally, Upper Paleolithic tools and artifacts communicate information such as status and group membership. Such innovation and social signaling seem to have been absent in Homo erectus, suggesting that they had a different relationship with their tools than did Homo sapiens (Coolidge and Wynn 2017). Some scientists assert that these contrasts in tool form and manufacture may signify key cognitive differences between the species, such as the ability to use a complex language.
Subsistence and Diet
In reconstructing the diet of Homo erectus, researchers can draw from multiple lines of evidence. These include stone tools used by Homo erectus, animal bones and occasionally plant remains from Homo erectus sites, and the bones and teeth of the fossils themselves. These data sources suggest that compared to the australopithecines, Homo erectus consumed more animal protein. Coinciding with the appearance of Homo erectus fossils in Africa are archaeological sites with much more abundant stone tools and larger concentrations of butchered animal bones.
Meat Eating and Increased Brain Size
It makes sense that a larger body and brain would be correlated with a dietary shift to more calorically dense foods. This is because the brain is a very energetically greedy organ. Indeed, our own human brains require more than 20% of one’s calorie total intake to maintain. When biologists consider the evolution of intelligence in any animal species, it is often framed as a cost/benefit analysis: In order for large brains to evolve, there has to be a compelling benefit to having them and a way to generate enough energy to fuel them.
One solution that would allow for an increase in human brain size would be a corresponding reduction in the size of the digestive tract (gut). According to the “expensive tissue hypothesis,” initially formulated by Leslie Aiello and Peter Wheeler (1995), a smaller gut would allow for a larger brain without the need for a corresponding increase in the organism’s metabolic rate. Judging from their skeleton, australopithecines have a wider rib cage and trunk region more similar to apes than humans. It is thought that the australopithecines had large gut sizes similar to today’s great apes because they were eating mainly plant foods, which require more gut bacteria to digest. More meat in the diet would allow for a smaller gut and could also fuel the larger brain and body size seen in the genus Homo. Some researchers also believe that body fat percentages increased in hominins (particularly females) around this time, which would have allowed them to be better buffered against environmental disruption such as food shortages (Anton and Snodgrass 2012).
Evidence for Dietary Versatility in Homo erectus
As indicated above, evidence from archaeology and the inferences about Homo erectus body size suggest increased meat eating. How much hunting did Homo erectus engage in compared to the earlier Oldowan toolmakers? Although experts continue to debate the relative importance of hunting versus scavenging, there seems to be stronger evidence of hunting for these hominins. For example, at sites such as Olorgesailie in Kenya (Figure \(\PageIndex{4}\), there are numerous associations of Acheulean tools with butchered remains of large animals.
 Figure \(\PageIndex{4}\): Excavations at the site of Olorgesailie, Kenya. Dated from between 1.2 million years ago and 490,000 years ago, Olorgesailie has some of the most abundant and well-preserved evidence of Homo erectus activity in the world. Fossils of large mammals, such as elephants, along with thousands of Acheulean tools, have been uncovered over the decades.
Figure \(\PageIndex{4}\): Excavations at the site of Olorgesailie, Kenya. Dated from between 1.2 million years ago and 490,000 years ago, Olorgesailie has some of the most abundant and well-preserved evidence of Homo erectus activity in the world. Fossils of large mammals, such as elephants, along with thousands of Acheulean tools, have been uncovered over the decades.However, Homo erectus certainly ate more than just meat. Modern-day hunter-gatherer societies have been used as models for considering the behavior patterns and environmental constraints of our early ancestors. Plant foods make up the bulk of calories for most modern-day hunter-gatherer societies, since they are a much more reliable food source. It would make sense that we would see similar patterns among early hominins.
Studies of the tooth surfaces and microscopic wear patterns on hominin teeth indicate that Homo erectus ate a variety of foods, including some hard, brittle plant foods (Unger and Scott 2009). This would make sense, considering the environment was changing to be more dominated by grasslands in some areas. Roots, bulbs, and tubers (known as underground storage organs) of open savanna plants may have been a primary food source. Indeed, hunter-gatherer groups such as the Hadza of Tanzania rely heavily on such foods, especially during periods when game is scarce. In the unstable environment of the early Pleistocene, dietary versatility would be a definite advantage.
Tool Use, Cooking, and Fire
One key characteristic of the genus Homo is smaller teeth compared to Australopithecus. Why would teeth get smaller? Besides the change in the type of foods eaten, there may have also been changes in how food was prepared and consumed. Think about how you would eat if you didn’t have access to cutting tools. What you couldn’t rip apart with your hands would have to be bitten off with your teeth—actions that would require bigger, more powerful teeth and jaws. During this time, stone tools were becoming increasingly important. If hominins were using these tools to cut up, tenderize, and process meat and plants, they wouldn’t have to use their teeth so vigorously.
Cooking food could also have contributed to the reduction in tooth and jaw size. In fact, anthropologist Richard Wrangham (2009) asserts that cooking played a crucial role in human evolution. Cooking provides a head start in the digestive process because of how heat begins to break down food before food even enters the body, and it can help the body extract more nutrients out of meat and plant foods such as starchy tubers. According to Wrangham’s model, this improved diet had a number of far-reaching consequences for human evolution. Most importantly, it allowed for the larger brain and body size (and smaller gut size) seen in Homo erectus.
Obviously cooking requires fire, and the earliest use of fire is a fascinating topic in the study of human evolution. Fire, of course is not limited to humans; it occurs naturally as a result of lightning strikes. Like other wild animals, early hominins must have been terrified of wildfires, but at some point in time learned to control fire and put it to good use. Cooking, warmth, and scaring off wild animals are just some of the benefits of fire. Consider the potential social benefits of having a light source after dark. Rather than just going to sleep, members of the group could repair tools, plan the next day’s activities, or socialize—just as you might do sitting around a campfire with family or friends. Isn’t it intriguing to think about how such activities might have encouraged the development of language?
Documenting the earliest evidence of fire has been a contentious issue in archaeology because of the difficulty in distinguishing between human-controlled fire and natural burning at hominin sites. Burned areas and ash deposits must have direct associations with human activity to make a case for deliberate fire use. Examples might include the presence of wood ash in caves where trees don’t naturally grow, deep ash deposits in hearths lined with stones, or burned pieces of stone tools and butchered animal bones (Gao 2017). Unfortunately, such evidence is rare at ancient hominin sites, which have been profoundly altered by humans, animals, and geological forces over millions of years. Recently, newer methods—including microscopic analysis of burned rock and bone—have revealed clear evidence of fire use at Koobi Fora, Kenya, dating to 1.5 million years ago (Hlubik et al. 2017).
MIGRATION OUT OF AFRICA
Homo erectus is generally thought to be the first hominin species to leave the continent of Africa and settle in Eurasia in places such as the Republic of Georgia, Indonesia, and northern China. We previously discussed the timing and fossil evidence for the appearance of Homo erectus at those sites; now we can address why the species traveled such vast distances to these far-flung regions. To do this, we have to consider what we have learned about the biology, culture, and environmental circumstances of Homo erectus. The larger brain and body size of Homo erectus were fueled by a diet consisting of more meat, and longer more powerful legs made it possible to walk and run longer distances to acquire food. Since they were eating higher on the food chain, it was necessary for them to extend their home range to find sufficient game. Cultural developments including better stone tools and new technology such as fire gave them greater flexibility in adapting to different environments. Finally, the major Pleistocene climate shift discussed earlier in the chapter certainly played a role. Changes in air temperature, precipitation, access to water sources, and other habitat alteration had far-reaching effects on animal and plant communities; this included Homo erectus. If hominins were relying more on hunting, the migration patterns of their prey could have led them increasingly long distances.
Life History
The life history of a species refers to its overall pattern of growth, development, and reproduction during its lifetime, with the assumption being that these characteristics have been shaped by natural selection. Our species, Homo sapiens, is characterized by a unique life history pattern of slow development, a long period of juvenile dependence, and a long lifespan. Unlike the great apes whose offspring achieve early self-sufficiency, human children are dependent on their parents long after weaning. Additionally, human fathers and grandparents (particularly post-menopausal grandmothers) devote substantial time and energy to caring for their children.
Human behavioral ecologists who study modern hunter-gatherer societies have observed that foraging is no easy business (Figure \(\pageindex{5}\). Members of these groups engage in complex foraging techniques that are difficult and take many years to master. An extended juvenile period gives children the time to acquire these skills. It also allows time for large human brains to grow and mature. On the back end, a longer developmental period results in skilled, successful adults, capable of living a long time (Hill and Kaplan 1999). Despite the time and energy demands, females could have offspring at more closely spaced intervals if they could depend on help from fathers and grandmothers (Hawkes et al. 1998).
Study of human behavior from an evolutionary and ecological perspective.
 Figure \(\PageIndex{5}\): Hadza men practice bowing. Native to Tanzania, the Hadza have retained many traditional foraging practices. Although most do not subsist entirely upon wild foods today, their way of life may shed light on how humans lived for most of their evolutionary history.
Figure \(\PageIndex{5}\): Hadza men practice bowing. Native to Tanzania, the Hadza have retained many traditional foraging practices. Although most do not subsist entirely upon wild foods today, their way of life may shed light on how humans lived for most of their evolutionary history.What can the study of Homo erectus reveal about its life history pattern? Well-preserved fossils such as the Nariokotome boy can provide some insights. We know that apes such as chimpanzees reach maturity more quickly than humans, and there is some evidence that the australopithecines had a growth rate more akin to that of chimpanzees. Scientists have conducted extensive studies of the Nariokotome skeleton’s bones and teeth to assess growth and development. On the one hand, examination of the long bone ends (epiphyses) of the skeleton suggested that he was an early adolescent with a relatively large body mass, though growth had not yet been completed. On the other hand, study of the dentition, including measurement of microscopic layers of tooth enamel called perikymata, revealed a much younger age of 8 or 9. According to Christopher Dean and Holly Smith (2009), the best explanation for this discrepancy between the dental and skeletal age is that Homo erectus had its own distinct growth pattern—reaching maturity more slowly than chimpanzees but faster than Homo sapiens. This suggests that the human life history pattern of slow maturation and lengthy dependency was a more recent development. More work remains on refining this pattern for early Homo, but it is an important question, as it sheds light on how and when we developed our unique life history characteristics (Table \(\PageIndex{2}\)).
Microscopic ridges on the surface of tooth enamel that serve as markers of tooth development.
|
Hominin |
Homo erectus |
|
Dates |
1.8 million years ago to about 200,000 years ago |
|
Region(s) |
East and South Africa; West Eurasia; China and Southeast Asia |
|
Famous Discoveries |
Lake Turkana, Olorgesailie, Kenya; Zhoukoudian, China; Dmanisi, Republic of Georgia |
|
Brain Size |
Average 900 cc; range between 650 cc and 1,100 cc |
|
Dentition |
Smaller teeth than Homo habilis |
|
Cranial Features |
Long, low skull with robust features including thick cranial vault bones and large brow ridge, sagittal keel, and occipital torus |
|
Postcranial Features |
Larger body size compared to Homo habilis; body proportions (longer legs and shorter arms) similar to Homo sapiens |
|
Culture |
Acheulean tools (in Africa); evidence of increased hunting and meat-eating; use of fire; migration out of Africa |
REFERENCES
Aiello, Leslie C., and Peter Wheeler. 1995. “The Expensive-Tissue Hypothesis.” Current Anthropology 36 (2): 199–221.
Anton, Susan C., and J. Josh Snodgrass. 2012. “Origins and Evolution of Genus Homo: New Perspectives.” Current Anthropology 53 (S6): S479–S496.
Ashton, Nick, Simon G. Lewis, Isabelle De Groote, Sarah M. Duffy, Martin Bates, Richard Bates, Peter Hoare, Mark Lewis, Simon A. Parfitt, Sylvia Peglar, Craig Williams, and Chris Stringer. 2014. “Hominin Footprints from Early Pleistocene Deposits at Happisburgh, UK.” PLOS ONE 9 (2): e88329.
Bramble, Dennis M., and Daniel E. Lieberman. 2004. “Endurance Running and the Evolution of Homo.” Nature 432: 345–352.
Coolidge, Frederick L., and Thomas Grant Wynn. 2017. The Rise of Homo Sapiens: The Evolution of Modern Thinking. New York: Oxford University Press.
Dean, M. Christopher, and B. Holly Smith. 2009. “Growth and Development of the Nariokotome Youth, KNM-WT 15000.” In The First Humans–Origin and Early Evolution of the Genus Homo: Contributions from the Third Stony Brook Human Evolution Symposium and Workshop October 3–7, 2006, edited by Frederick E. Grine, John G. Fleagle, and Richard E. Leakey, 101–120. Dordrecht: Springer Netherlands.
Gao, Xing, Shuangquan Zhang, Yue Zhang, and Fuyou Chen. 2017. “Evidence of Hominin Use and Maintenance of Fire at Zhoukoudian.” Current Anthropology 58 (Number S16): S267–S277.
Hawkes, Kristen, James F. O’Connell, Nicholas G. Blurton Jones, Helen Alvarez, and Eric L. Charnov. 1998. “Grandmothering, Menopause, and the Evolution of Human Life Histories.” Proceedings of the National Academy of Sciences 95 (3): 1336–1339.
Hill, Kim, and Hillard Kaplan. 1999. “Life History Traits in Humans: Theory and Empirical Studies.” Annual Review of Anthropology 28 (1): 397–430.
Hlubik, Sarah, Francesco Berna, Craig Feibel, David Braun, and John W. K. Harris. 2017. “Researching the Nature of Fire at 1.5 Mya on the Site of FxJj20 AB, Koobi Fora, Kenya, Using High-Resolution Spatial Analysis and FTIR Spectrometry.” Current Anthropology 58 (S16): S243–S257.
Jablonski, Nina G. 2010. “The Naked Truth.” Scientific American 302 (2): 42–49.
Kamberov, Yana G., Elinor K. Karlsson, Gerda L. Kamberova, Daniel E. Lieberman, Pardis C. Sabeti, Bruce A. Morgan, and Clifford J. Tabin. 2015. “A Genetic Basis of Variation in Eccrine Sweat Gland and Hair Follicle Density.” Proceedings of the National Academy of Sciences 112 (32): 9932–9937.
Kittler, R., M. Kayser, and M. Stoneking. 2003. “Molecular Evolution of Pediculus Humanus and the Origin of Clothing.” Current Biology 13 (16): 1414-7.
Lordkipanidze, David, Marcia S. Ponce de León, Ann Margvelashvili, Yoel Rak, G. Philip Rightmire, Abesalom Vekua, and Christoph P. E. Zollikofer. 2013. “A Complete Skull from Dmanisi, Georgia, and the Evolutionary Biology of Early Homo.” Science 342 (6156): 326–333.
Qiu, Jane. 2016. “How China Is Rewriting the Book on Human Origins.” Nature 535: 22-25.
Reed, David L., Jessica E. Light, Julie M. Allen, and Jeremy J. Kirchman. 2007. “Pair of Lice Lost or Parasites Regained: The Evolutionary History of Anthropoid Primate Lice.” BMC Biology 5 (1):7. doi: 10.1186/1741-7007-5-7.
Shipton, Ceri, James Blinkhorn, Paul S. Breeze, Patrick Cuthbertson, Nick Drake, Huw S. Groucutt, Richard P. Jennings, Ash Parton, Eleanor M. L. Scerri, Abdullah Alsharekh, and Michael D. Petraglia. 2018. “Acheulean Technology and Landscape Use at Dawadmi, Central Arabia.” PloS one 13 (7): e0200497–e0200497.
Simpson, Scott W., Jay Quade, Naomi E. Levin, Robert Butler, Guillaume Dupont-Nivet, Melanie Everett, and Sileshi Semaw. 2008. “A Female Homo erectus Pelvis from Gona, Ethiopia.” Science 322 (5904): 1089–1092.
Ungar, Peter S and Robert S. Scott. 2009. “Dental Evidence for Diets of Early Homo.” In The First Humans: Origin and Early Evolution of the Genus Homo, edited by Frederick E. Grine, John G. Fleagle, and Richard E. Leakey, 121-134. New York: Springer.
Wrangham, Richard. 2009. Catching Fire: How Cooking Made Us Human. New York: Basic Books.
Zhu, Zhaoyu, Robin Dennell, Weiwen Huang, Yi Wu, Shifan Qiu, Shixia Yang, and Zhiguo Rao. 2018. “Hominin Occupation of the Chinese Loess Plateau Since about 2.1 Million Years Ago.” Nature 559: 608–612.
FIGURE ATTRIBUTIONS
Figure \(\PageIndex{1}\): Homo erectus: Sangiran 17 lateral left view by eFossils is copyrighted and used for non-commercial purposes as outlined by eFossils.
Figure \(\PageIndex{2}\) Homo erectus site map original to Explorations: An Open Invitation to Biological Anthropology by Chelsea Barron at GeoPlace, California State University, Chico is under a CC BY-NC 4.0 License.
Figure \(\PageIndex{3}\) Hand axe spanish by Locutus Borg has been designated to the public domain (CC0).
Figure \(\PageIndex{4}\) Elephant Butchery Site Olorgesailie, Kenya by Smithsonian [exhibit: Human Evolution Research, East African Research Projects, Olorgesailie, Kenya] is copyrighted and used for educational and non-commercial purposes as outlined by the Smithsonian.
Figure \(\PageIndex{5}\) Hadzabe1 by Idobi is used under a CC BY-SA 3.0 License.
TABLE ATTRIBUTIONS
Table \(\PageIndex{1}\) Regional comparisons of Homo erectus fossils original to Explorations: An Open Invitation to Biological Anthropology is under a CC BY-NC 4.0 License.
Table \(\PageIndex{2}\) Summary features of Homo erectus original to Explorations: An Open Invitation to Biological Anthropology is under a CC BY-NC 4.0 License.