
7.5: Touch
- Page ID
- 173139
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)1. Describe the role of social and affective touch in development and bonding.
2. Describe how the somatosensory system allows for the sensing of touch, temperature, and position in space.
3. Describe the process of transduction in the senses of touch and proprioception.
4. Explain how expectations and context affect pain and touch experiences.
Overview
The somatosensory system is one of the most varied sensory systems on just about every level from initial types of stimuli to the wide range of ultimate perceptions.This system is regulated by specialized receptors that are spread throughout the body and measure a number of different sensory modalities in the body. These sensations can be divided into three main divisions: external stimuli, internal stimuli, and the sense of where the body is in space.
Perception of external stimuli can include the sense of touch (via mechanoreceptors), pain (via nociceptors), and temperature (via thermal receptors). Perception of internal stimuli can include organ (visceral) sensation and pain (via multiple receptor types) and blood chemical composition (via chemoreceptors). Finally, proprioception (via proprioceptors) is the sense of where the body is in space. The ability of an individual to touch their nose easily while their eyes are closed is an example of the proprioception system.
This section will provide an overview of most of these types of somatosensory inputs and the neural mechanisms that govern them.
Somatosensory Cell Bodies
All somatosensory receptor neurons have their cell bodies located in the dorsal root ganglion, a structure found just outside the dorsal aspect of the spinal cord (See Figure \(\PageIndex{1}\)). The receptor neurons, also called primary afferent fibers, of the somatosensory system are bipolar neurons, meaning they have one process from the cell body that splits into two branches. One travels to the location of the receptor (e.g. the skin for touch) via the spinal nerves, and one travels into the spinal cord at the dorsal horn via the dorsal root. The axon can either synapse in the spinal cord or ascend to the brain in the dorsal column.

Touch
The sense of touch is one of the most important senses in human development, particularly with regard to attachment to others. It has long been viewed as an essential part of childhood development (Baysinger, Plubell, & Harlow, 1973; Feldman, 2007; Haradon, Bascom, Dragomir, & Scripcaru, 1994) and also helps us feel socially attached to others (Field et al., 1997; Kelter, 2009). Touch is an essential part of everyday activities ranging from touching our food to eat to sharing human affection with another individual. Recent studies indicate that slow affectionate touch, rather than faster neutral touch, can serve as a buffer against physical pain. In addition, the same slow affectionate touch can reduce feelings of social exclusion and increase social bonding.
Skin is the largest organ in our body and has a variety of nerve endings that allow us to feel sensations such as pressure, pain, cold, and heat. The skin on our bodies also serves as a protector to important vital organs.
Touch and pain are aspects of the somatosensory system, which provides our brain with information about our own body (interoception) and properties of the immediate external world (exteroception) (Craig, 2002). We have somatosensory receptors located all over the body, from the surface of our skin to the depth of our joints. As indicated previously, the information they send to the central nervous system is generally divided into four modalities: cutaneous senses (senses of the skin), proprioception (body position), kinesthesis (body movement), and nociception (pain, discomfort).
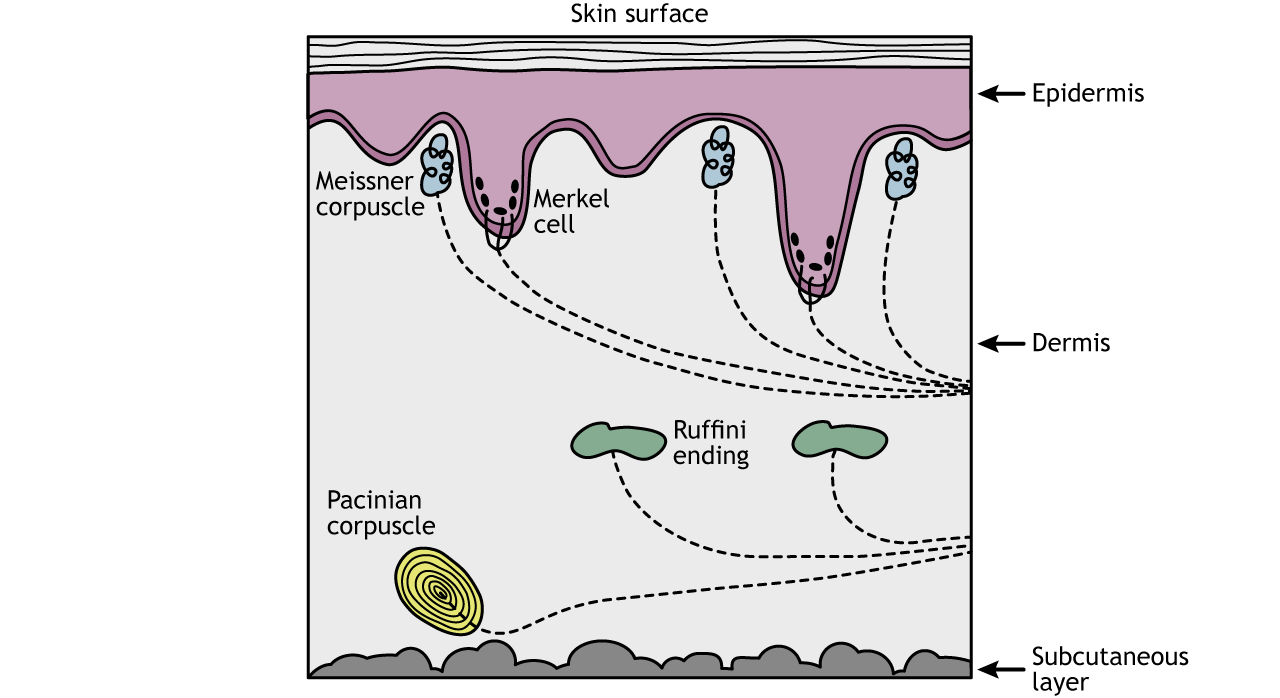
Cutaneous Senses (senses of the skin)
Cutaneous senses respond to tactile, thermal, and pruritic (itchy) stimuli, and events that cause tissue damage (pain). Cutaneous mechanoreceptors are located in the skin. They provide the senses of touch, pressure, vibration, proprioception, and others. They all utilize nerves known as Aβ fibers from the skin that are mostly dedicated to touch, except for the mechano-receiving free nerve endings, which use Aδ fibers to process information primarily relating to acute pain; sharp, immediate, and relatively short-lasting.
The can be categorized by morphology, by the type of sensation they perceive, and by the rate of adaptation. Furthermore, each has a different receptive field:
- Ruffini’s end organs detect tension deep in the skin.
- Meissner’s corpuscles detect changes in texture (vibrations around 50 Hz) and adapt rapidly.
- Pacinian corpuscles detect rapid vibrations (about 200–300 Hz).
- Merkel’s discs detect sustained touch and pressure.
- Mechanoreceiving free nerve endings detect touch, pressure, and stretching.
- Hair follicle receptors are located in hair follicles and sense the position changes of hair strands.
Ruffini Ending
The Ruffini ending (Ruffini corpuscle or bulbous corpuscle) is a class of slowly adapting mechanoreceptors thought to exist only in the glabrous dermis and subcutaneous tissue of humans. It is named after Angelo Ruffini (an Italian histologist and embryologist).
This spindle-shaped receptor is sensitive to skin stretch, and contributes to the kinesthetic sense of and control of finger position and movement. It is believed to be useful for monitoring the slippage of objects along the surface of the skin, allowing the modulation of grip on an object.
Ruffini endings are located in the deep layers of the skin. They register mechanical information within joints, more specifically angle change, with a specificity of up to two degrees, as well as continuous pressure states. They also act as thermoreceptors that respond for a long time, such as holding hands with someone during a walk. In a case of a deep burn to the body, there will be no pain as these receptors will be burned off.
Meissner’s Corpuscles
Meissner’s corpuscles (or tactile corpuscles) are responsible for sensitivity to light touch. In particular, they have the highest sensitivity (lowest threshold) when sensing vibrations lower than 50 hertz. They are rapidly adaptive receptors.
Pacinian Corpuscles
Pacinian corpuscles (or lamellar corpuscles) are responsible for sensitivity to vibration and pressure. The vibrational role may be used to detect surface texture, e.g., rough versus smooth.
Merkel Nerve
Merkel nerve endings are mechanoreceptors found in the skin and mucosa of vertebrates that provide touch information to the brain. The information they provide are those regarding pressure and texture. Each ending consists of a Merkel cell in close apposition with an enlarged nerve terminal.
This is sometimes referred to as a Merkel cell–neurite complex, or a Merkel disk receptor. A single afferent nerve fiber branches to innervate up to 90 such endings. They are classified as slowly adapting type I mechanoreceptors.
Proprioception (body position)
Proprioception refers to the sense of knowing how one’s body is positioned in three-dimensional space. Proprioception is the sense of the relative position of neighboring parts of the body and the strength of effort being employed in movement. It is distinguished from exteroception (perception of the outside world) and interoception (perception of pain, hunger, and the movement of internal organs). The initiation of proprioception is the activation of a proprioreceptor in the periphery. The proprioceptive sense is believed to be composed of information from sensory neurons located in the inner ear (motion and orientation) and in the stretch receptors located in the muscles and the joint-supporting ligaments (stance). Conscious proprioception is communicated by the posterior (dorsal) column–medial lemniscus pathway to the cerebrum. Unconscious proprioception is communicated primarily via the dorsal and ventral spinocerebellar tracts to the cerebellum.
An unconscious reaction is seen in the human proprioceptive reflex, or Law of Righting, a reflex that corrects the orientation of the body when it is taken out of its normal upright position. In the event that the body tilts in any direction, the person will tilt their head back to level the eyes against the horizon. Infants are often seen doing this as soon as they gain control of their neck muscles. This control comes from the cerebellum, the part of the brain that affects balance.
Muscle spindles are sensory receptors within the belly of a muscle that primarily detect changes in the length of a muscle. They convey length information to the central nervous system via sensory neurons. This information can be processed by the brain to determine the position of body parts. The responses of muscle spindles to changes in length also play an important role in regulating the contraction of muscles.
The Golgi organ (also called Golgi tendon organ, tendon organ, neurotendinous organ or neurotendinous spindle) is a proprioceptive sensory receptor that provides the sensory component of the Golgi tendon reflex. The Golgi organ should not be confused with the Golgi apparatus—an organelle in the eukaryotic cell —or the Golgi stain, which is a histologic stain for neuron cell bodies. The Golgi tendon reflex is a normal component of the reflex arc of the peripheral nervous system. In a Golgi tendon reflex, skeletal muscle contraction causes the agonist muscle to simultaneously lengthen and relax. This reflex is also called the inverse myotatic reflex, because it is the inverse of the stretch reflex. Although muscle tension is increasing during the contraction, alpha motor neurons in the spinal cord that supply the muscle are inhibited and antagonistic muscles are activated.
Kinesthesis (body movement)
Kinesthesis or Kinesthesia is a term that is often used interchangeably with proprioception. Some users differentiate the kinesthetic sense from proprioception by excluding the sense of equilibrium or balance from kinesthesia. An inner ear infection, for example, might degrade the sense of balance. This would degrade the proprioceptive sense, but not the kinesthetic sense. The infected person would be able to walk, but only by using their sense of sight to maintain balance; the person may have difficulty walking when their eyes are closed.
Proprioception and kinesthesia are seen as interrelated and there is considerable disagreement regarding the definition of these terms. Some of this difficulty stems from Sherrington's original description of joint position sense (or the ability to determine where a particular body part exactly is in space) and kinesthesia (or the sensation that the body part has moved) under a more general heading of proprioception. Clinical aspects of proprioception are measured in tests that measure a subject's ability to detect an externally imposed passive movement, or the ability to reposition a joint to a predetermined position. Often it is assumed that the ability of one of these aspects will be related to another, unfortunately experimental evidence suggests there is no strong relation between these two aspects. This suggests that while these components may well be related in a cognitive manner, they seem to be separate physiologically.
Much of the forgoing work is dependent on the notion that proprioception is essentially a feedback mechanism: that is the body moves (or is moved) and then the information about this is returned to the brain whereby subsequent adjustments could be made. More recent work into the mechanism of ankle sprains suggest that the role of reflexes may be more limited due to their long latencies (even at the spinal cord level) as ankle sprain events occur in perhaps 100 milliseconds or less. As a result, a model has been proposed to include a 'feedforward' component of proprioception where the subject will also have central information about the body's position prior to attaining it.
Kinesthesia is a key component in muscle memory and hand-eye coordination and training can improve this sense. The ability to swing a golf club, or to catch a ball, requires a finely-tuned sense of the position of the joints. This sense needs to become automatic through training to enable a person to concentrate on other aspects of performance, such as maintaining motivation or seeing where other people are.
Nociception (pain, discomfort)
Nociception deals with a series of events and processes required for an organism to receive a painful stimulus, convert it to a molecular signal, and recognize and characterize the signal in order to trigger an appropriate defense response. This serves as an important function because our brains can often turn off or reduce the feelings of pain depending on the situation we may be experiencing or involved in. An example involves athletes who often do not feel pain until after they have completed the game or event they are participating in (Bantick, Wise, Ploghaus, Clare, Smith, & Tracey, 2002). In essence their brain is involved in a complicated process that requires them to utilize multiple systems to stay focused on the activity. The endorphins that are released when we are engaged or excited by a situation or event around us act as natural pain killers (Sternberg, Bailin, Grant, & Gracely, 1998).
Spinal Pathways: From the Body to the Brain
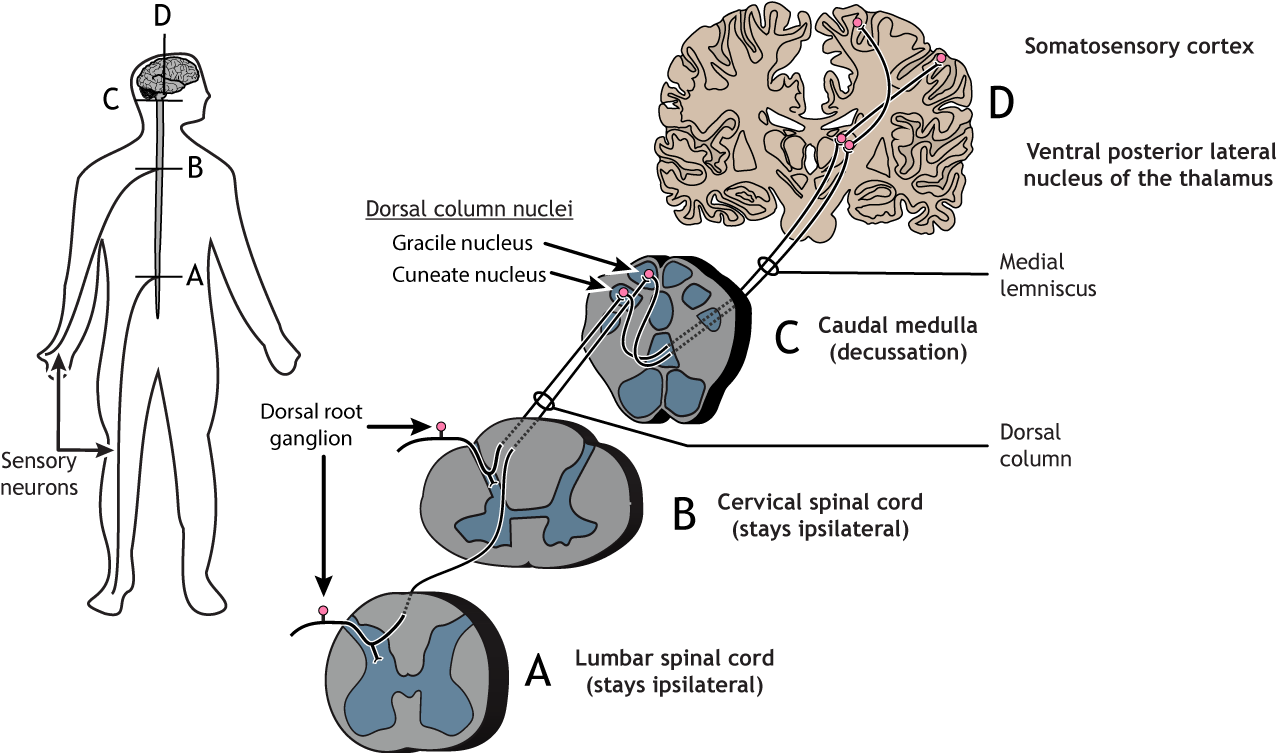
All of the various types of somatosensory input described above must make its way up to cortical areas of the brain to ultimately be perceived and acted upon. As mention in the cutaneous section, there is may be multiple pathways that an input can take depending on the nature of the stimuli being processes. These incoming pathways are often referred to as afferent axons to denote that the information will ultimately travel via axons making their way into the central nervous system.
Primary afferent axons are divided into four groups based on size and conduction speed. The groups, unfortunately, have different names depending on if the axons come from the skin (Aα, Aβ, Aδ, and C fibers; examples are touch or pain) or the muscles (Group I, II, III and IV fibers; example is proprioception). The fastest axons are the Aα or Group I type; they have the largest diameter and are heavily myelinated. The C fibers have the smallest diameter, are unmyelinated, and are the slowest at conducting action potentials. See Figure \(\PageIndex{3}\) for an illustration of how these fibers enter the nervous and make their way up to the brain. Note that the axons cross to the opposite side at some point, so the cortex ultimately processes information from the contralateral (opposite) side of the body.
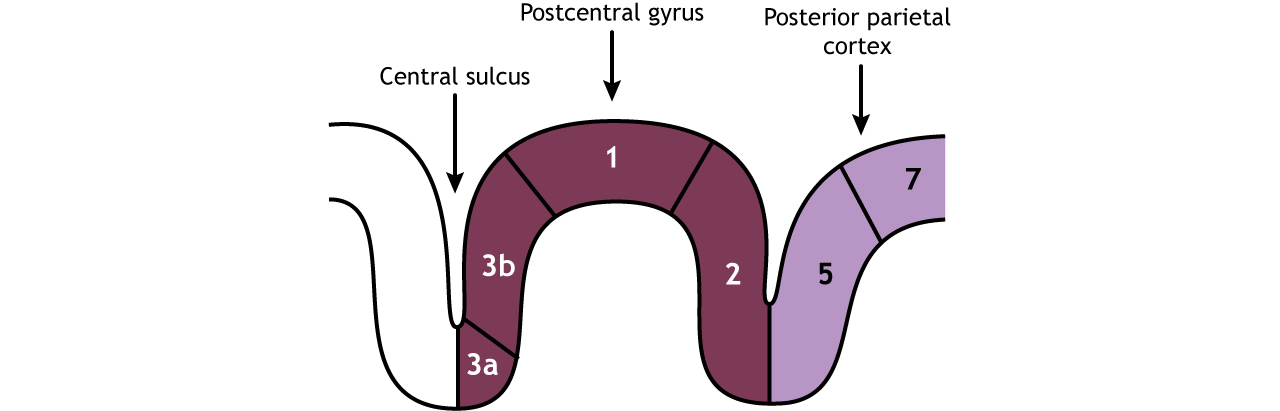
The Primary Somatosensory Cortex
The primary somatosensory cortex is divided into four regions, each with its own input and function: areas 1, 2, 3a, and 3b. Most touch information from mechanoreceptors inputs to region 3b, whereas most proprioceptive information from the muscles inputs to region 3a. These regions then send and receive information from areas 1 and 2. As processing of somatosensory information continues, the stimuli required to activate neurons becomes more complex. For example, area 1 is involved in sensing texture, and area 2 is involved in sensing size and shape of an object. (see Figure \(\PageIndex{4}\)).
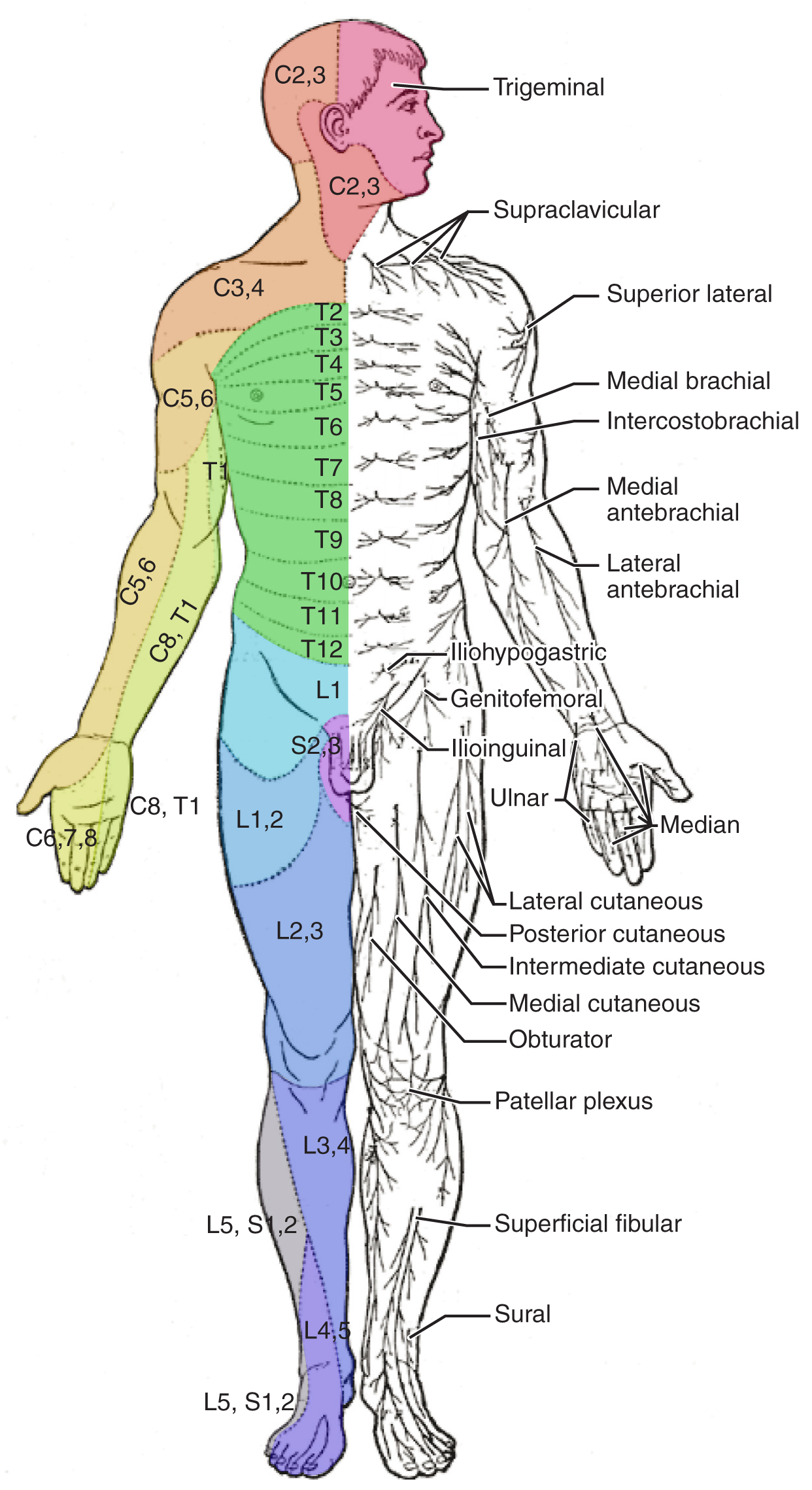
Testing of the senses begins with examining the regions known as dermatomes that connect to the cortical region where somatosensation is perceived in the postcentral gyrus. To test the sensory fields, a simple stimulus of the light touch of the soft end of a cotton-tipped applicator is applied at various locations on the skin. The spinal nerves, which contain sensory fibers with dendritic endings in the skin, connect with the skin in a topographically organized manner, illustrated as dermatomes (Figure \(\PageIndex{5}\) - it is not necessary to memorize the terms in this figure). For example, the fibers of the eighth cervical nerve innervate the medial surface of the forearm and extend out to the fingers. In addition to testing perception at different positions on the skin, it is necessary to test sensory perception within the dermatome from distal to proximal locations in the appendages, or lateral to medial locations in the trunk. In testing the eighth cervical nerve, the patient would be asked if the touch of the cotton to the fingers or the medial forearm was perceptible, and whether there were any differences in the sensations.
Attributions
- Touch sections adapted by Isaias Hernandez from "NOBA-Biological Psychology, Touch and Pain"; https://nobaproject.com/modules/touch-and-pain#:~:text=Touch%20and%20pain%20are%20aspects,)%20(Craig%2C%202002).
- Touch sections adapted by Isaias Hernandez from "Introduction to Psychology, University of Minnesota"'; https://open.lib.umn.edu/intropsyc/chapter/4-4-tasting-smelling-and-touching/
- Touch sections adapted by Isaias Hernandez from "Boundless Anatomy and Physiology from Lumen"; https://courses.lumenlearning.com/boundless-ap/chapter/pain/
- Touch sections adapted by Isaias Hernandez from "Kinesthia, Psychology Wiki"; https://en.Wikipedia.org/wiki/Kinaesthetics
- Touch sections adapted by Isaias Hernandez from "Nociception, Wikipedia"; https://en.Wikipedia.org/wiki/Nociception
- Touch sections adapted by Isaias Hernandez from "Foundations of Neuroscience, Michigan State University"; https://openbooks.lib.msu.edu/neuroscience/chapter/touch-the-skin/
- Touch sections adapted by Isaias Hernandez from "Anatomy and Physiology, Oregon State University"; https://open.oregonstate.education/aandp/chapter/14-5-sensory-and-motor-pathways/
- Further revision of most sections by Alan Keys, Ph.D., Sacramento City College, Sacramento, CA.
References
- von Mohr, M., Kirsch, L.P. & Fotopoulou, A. The soothing function of touch: affective touch reduces feelings of social exclusion. Sci Rep 7, 13516 (2017). https://doi.org/10.1038/s41598-017-13355-7