
5.2: Within-neuron Communication- Electrical Potentials from Resting to Action
- Page ID
- 173127
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Overview
Information processing that contributes to the workings of our minds and control behavior depend upon neurons and their electrical and chemical interactions. Neurons produce electrical potentials (voltages) that act as signals in the information processing activities of the brain. As previously mentioned, neurons communicate with one another across synaptic spaces using chemicals known as neurotransmitters. However, before communication can take place between neurons, there is a series of electrical changes that must occur within neurons. In this module, we examine how neurons create these electrical changes or potentials including the resting, post-synaptic, and action potential.
Within-Neuron Potentials: How Neurons Produce Electrical Potentials
Before a neuron can send a chemical message to another neuron, muscle, or gland it usually must experience an electrical change within itself. Neurons are capable of producing electrical activity (i.e. voltage potentials) by the unequal distribution and movement of electrically charged atoms called ions across the neuron's cell membrane. These ions come mainly from dissolved salts in the body fluids inside and outside neurons.
The main ions used by neurons to produce their voltages are sodium (Na+), potassium (K+), and chloride (Cl-). Notice that the first two are positive ions and the last, chloride ions, are negatively charged. A fourth ion, organic anions (A-), which are large (on the molecular scale) negatively charged proteins, are manufactured inside the neuron. They are too large to cross the cell membrane, and therefore give the neuron's resting voltage a negative bias. The distribution or concentrations of these ions inside and outside a neuron determine its voltage (voltage is just the physical separation of charged particles, like in a car battery with its positive and negative poles around which are concentrated positively and negatively charged particles floating in battery acid).
By these means, three main types of neuron voltages or neuron potentials are produced by neurons: 1) the resting potential (often equal to about negative 70 thousandths of a volt, -70 millivolts) 2) post-synaptic potentials (PSPs)-- which can be excitatory (EPSPs) or inhibitory (IPSPs) and 3) the action potential (AP, the nerve impulse). The action potential is the last step of within-neuron communication which then triggers the chemical communication between neurons. Let's examine these potentials in more detail and see how they are generated inside neurons.
The Neuronal Cell Membrane
The neuronal cell membrane, which is composed of a lipid bilayer of fat molecules, separates the fluid inside of the cell from the surrounding extracellular fluid. There are proteins that span the membrane, forming ion channels that allow, when open, particular ions to pass between the intracellular and extracellular fluid (see Figure \(\PageIndex{1}\)). These ions are in different concentrations inside the cell relative to outside the cell, and the ions have different electrical charges. Due to this difference in ion concentration and charge, in part enforced by the physical barrier of the cell membrane when ion channels to specific ions are closed, a voltage is produced, the resting potential.
The Resting Membrane Potential
The resting potential can be thought of as a baseline voltage from which the other neuron potentials are generated. It is the voltage inside a nerve cell when it is at rest, that is, it is neither receiving inputs at the moment nor generating any outputs. In this state of "rest", the voltage inside the nerve cell is approximately -70 mv. It is negative inside the neuron at "rest" because there are more negative ions inside the neuron (the intracellular fluid) and more positive ions on the outside (the extracellular fluid). Specifically, there are large numbers of Na+ ions (sodium ions) outside the neuron and very few of these on the inside, when the neuron is "at rest". And there are more negatively charged ions inside the cell than on the outside during resting potential. During the resting potential ion channels to Na+ are closed.
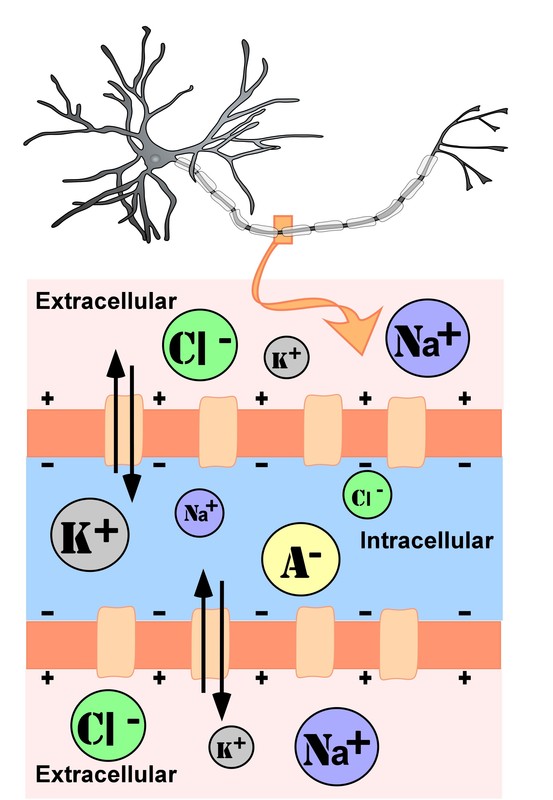
Figure \(\PageIndex{1}\): Representation of ion concentrations inside (intracellular) and outside (extracellular) a neuron at rest in the unmyelinated segment of the axon. Size of the circles represents relative concentrations of each ion inside and outside the neuron; note that when the neuron is "at rest" it has a net negative charge inside, the resting potential, equal to about -70 millivolts in most neurons. Also note the higher concentrations of sodium (Na+) and chloride (Cl-) ions outside the cell. Sodium on the outside of the neuron "would like" to move into the cell along its charge and concentration gradients, but it can't when the cell is at rest, because the ion channels for sodium ions are closed, creating a physical barrier keeping most of the extracellular sodium ions from crossing through the cell membrane. But what happens when the sodium channels open? And, test yourself, what opens the ion channels? (Authored by: R. Biswas-Diener & E. Diener (Eds), Noba textbook series: Psychology. Champaign, IL: DEF publishers, Neurons by Sharon Furtak at NOBA is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License.
The Sodium-Potassium Pump
During the resting potential, the sodium-potassium pump maintains a difference in charge across the cell membrane of the neuron. The sodium-potassium pump is a mechanism of active transport that moves sodium ions out of cells and potassium ions into cells. The sodium-potassium pump moves both ions from areas of lower to higher concentration, using energy in ATP and carrier proteins in the cell membrane (see Figure \(\PageIndex{2}\)).
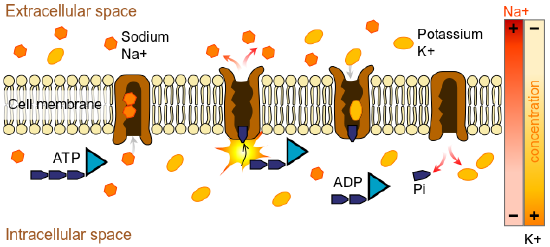
Figure \(\PageIndex{2}\): The sodium-potassium pump helps maintain the resting potential of a neuron. During resting potential, there is more negative charge inside than outside the cell creating a resting potential of -70mv. During resting potential, some Na+ leaks into the neuron and some K+ leaks out. ATP (Adenosine triphosphate) provides energy to pump sodium out and potassium into the cell. There is more concentration of sodium outside the membrane and more concentration of potassium inside the cell due in part to the unequal movement of these ions by the pump. The presence of negatively charged organic anions (A-) contributes to the net negative charge (-70mv) inside the neuron at rest. (Image from Wikimedia Commons; File:Scheme sodium-potassium pump-en.svg; https://commons.wikimedia.org/wiki/F...um_pump-en.svg; by LadyofHats Mariana Ruiz Villarreal; released into the public domain by its author, LadyofHats. This applies worldwide).
Electrical and Concentration Gradients: How Ions Move Across the Neuronal Membrane.
For a neuron to move away from the resting state, other neuron potentials need to be generated. In other words, a redistribution of ions (and their electrical charges) must occur across the cell membrane resulting in a positive or negative voltage change in the neuron. In short, ions must move.
There are two main forces (called gradients) that can cause these ions to move.
First, opposite charges attract one another ("opposites attract"), and like charges repel one another. When ions of opposite charge are unequally distributed across the cell membrane (as is the case during the resting potential), this sets up what is called a charge or electrical gradient (an unequal distribution of charged particles).This gradient then creates an electrostatic force. If these ions are allowed to move freely, they will move along this electrical gradient with positive charges moving toward negative ones and vice versa.
Secondly, when ions of any particular type (for example, sodium ions) are unequally distributed across the cell membrane (like during the resting potential), this sets up what is called a concentration gradient which can cause diffusion. Diffusion is when the net movement of ions from a region of higher concentration to a region of lower concentration occurs. Such concentration gradients for several ions (sodium, potassium, and chloride ions) exist when the neuron is at resting potential (see Figure \(\PageIndex{1}\)). Ions, if allowed to move freely, will move along their concentration gradients, such that ions of a particular type (like Na+ ions) will move from a region of high concentration (of Na+) to a region of lower concentration (of Na+).
In short, ions, if allowed to move freely, will move along both their electrical and concentration gradients. Ions "want" to move to equalize their concentrations across the cell membrane and also "want" to move to equalize the charges across the cell membrane.
However, it is important to note that these two forces created by the electrical and concentration gradients for each ion can oppose one another in the case of some ions, or can act together (as is the case with Na+ ions during the resting potential).
Let us see how these two forces, diffusion (due to concentration gradients) and electrostatic pressure (due to a charge gradient), act on the four groups of ions mentioned above.
- Anions (A-): Anions are highly concentrated inside the cell and contribute to the negative charge of the resting membrane potential. Diffusion and electrostatic pressure are not forces that determine A- concentration because A- is impermeable to the cell membrane. There are no ion channels that allow for A- to move between the intracellular and extracellular fluid.
- Potassium (K+): The cell membrane is very permeable to potassium at rest, but potassium remains in high concentrations inside the cell. Diffusion created by the concentration gradient pushes K+ to the outside of the cell because it is in high concentration inside the cell. However, electrostatic pressure created by the charge gradient pushes K+ into the cell because the positive charge of K+ is attracted to the negative charge inside the cell. In combination, these forces oppose one another with respect to K+ with the charge gradient (opposite charges attract) overpowering the concentration gradient so that the net effect is to push and keep K+ inside the neuron in higher concentrations.
- Chloride (Cl-): The cell membrane is also very permeable to chloride at rest, but chloride remains in high concentration outside the cell. Diffusion created by a concentration gradient for Cl- pushes Cl- toward the inside of the cell because it is in high concentration outside the cell. However, electrostatic pressure created by the charge gradient for Cl- pushes Cl- toward the outside of the cell because the negative charge of Cl- is attracted to the positive charge outside the cell created primarily by the high concentration of Na+ there. Similar to K+, these forces oppose one another with respect to Cl- and again, in this case, like Na+, the more powerful charge gradient (like charges repel) overcomes the weaker concentration gradient for Cl-.
- Sodium (Na+): The cell membrane is not very permeable to sodium when the neuron is at rest. Diffusion created by a concentration gradient pushes Na+ toward the inside of the cell because it is in high concentration outside the cell. Electrostatic pressure created by the charge gradient for Na+ also pushes Na+ toward the inside of the cell because the positive charge of Na+ is attracted to the negative charge inside the cell. Both of these forces push Na+ inside the cell; however, as discussed above, Na+ cannot permeate the cell membrane because the channels for Na+ are closed and so Na+ remains in high concentration outside the cell. The small amounts of Na+ inside the cell are removed by the sodium-potassium pump (Figure 5.2.2), which uses the neuron’s energy (adenosine triphosphate, ATP) to pump three Na+ ions out of the cell in exchange for bringing two K+ ions inside the cell.
Though ions "want" to move in these ways dictated by their charge gradients and concentration gradients, Na+ can't move freely when the neuron is at resting membrane potential because Na+ ion channels are closed. Thus, the cell membrane acts as a physical barrier to Na+ ions, preserving the unequal distributions of Na+ ions inside and outside the neuron. To get ion movement, pores or ion channels in the cell membrane of the neuron "at rest" must be opened, overcoming the physical barrier created by the cell membrane when the neuron is "at rest" (not receiving any inputs via its dendrites and not generating any outputs, action potentials, via its axon) and its sodium channels are closed.
What is it that causes the ion channels to open allowing the ions to move along their gradients? What is it that opens the ion channels? In reality, there are several possible triggers for ion channel changes, many of which are related to sensory input (physical stimulation to dendrites, chemical input from taste or smell systems, light or sound energy, etc.) However, the axonal release and subsequent arrival of transmitter molecules onto the postsynaptic neuron is the main trigger for PSPs that is presented here. In short, the "lock and key" interaction between molecules of transmitter and the post-synaptic receptor sites causes "doors"(specific ion channels) to open.
When specific ion channels get opened in this way (Na+ channels, for example), those specific ions move through the cell membrane along their specific concentration and electrical gradients. This will cause a voltage shift away from resting potential. This voltage shift is the post-synaptic potential. n this case it is an excitatory post-synaptic potential (EPSP), a positive shift in voltage away from resting potential because when Na+ channels in the cell membrane are opened by neurotransmitter, then Na+ ions will move along their charge gradient and concentration gradient into the cell, making the inside of the neuron more positively charged.
Excitatory and Inhibitory Post-synaptic Potentials (EPSPs and IPSPs)
As mentioned above, the cell membrane of a neuron has channels or "doors" for ions (electrically charged atoms) which can pass through the membrane when specific channels are opened for specific ions. Normally the ion channels on the membrane of a postsynaptic neuron are closed until they experience a sensory input or are acted upon by the attachment of neurotransmitter molecules to receptors sites on the membrane. The attachment of the molecules of transmitter to the receptors sites (receptor binding), like a key into a lock, opens the "doors" (ion channels) to specific ions which pass through the cell membrane carrying with them their electrical charge.
This ion channel opening results in a change in the electrical state of the neuron, a post-synaptic potential (i.e. a voltage shift in the post-synaptic neuron). If a positive shift in voltage occurs, it is called an EPSP, an excitatory post-synaptic potential; if a negative shift in voltage occurs, then it is called an IPSP, an inhibitory post-synaptic potential. EPSPs are triggered when neurotransmitter binds to "excitatory" receptors, and IPSPs are produced when transmitter binds to "inhibitory" receptors. Receptors are classified as excitatory if their transmitter binding results in an excitation or increase in voltage of the neuron. They are classified as inhibitory if they result in the decrease of the voltage of the neuron.
These PSPs, whether excitatory or inhibitory, are called graded potentials because they are not of a fixed voltage, but instead vary in voltage depending on the amount of neurotransmitter (and other factors) that has been released into the synapse. This is in contrast to the action potential (the nerve impulse) which is of a fixed voltage and is "all or none," The all or none law will be fully detailed in the action potential section below. Think of two types of light switches: a typical on/off switch vs. a dimmer switch. A typical on/off switch only has two options and is similar to the all or none action potential, whereas, the dimmer switch has multiple options along a continuum and is most similar to the graded PSPs.
These post-synaptic potentials occur in a post-synaptic neuron (thus the name), a receiver neuron. When such a neuron receives an input from a pre-synaptic neuron, in the form of transmitter molecules which attach to the post-synaptic receptor sites (which are proteins that are embedded in the membrane of the post-synaptic cell), ion channels then open, ions move across the cell membrane, carrying with them their electrical charges. These events change the voltage inside the post-synaptic neuron. It is this voltage shift away from resting potential that constitutes the post-synaptic potential (PSP).
Note, again, that in both the EPSP and the IPSP, it is the attachment of molecules of transmitter to post-synaptic receptor sites (like keys going into locks of a specific shape) that opens the "doors", the ion channels, allowing ions to move through the cell membrane.
But there is one more key issue here. What is it that determines which ion channels open, and therefore, whether an EPSP or an IPSP occurs? The answer was implied above. The answer is: it is the type of neurotransmitter and type of receptor site receiving the transmitter molecules that determines the ultimate effect.
Excitatory transmitters (such as glutamate, acetylcholine--ACh, norepinephrine--NE, dopamine--DA) are those which open positive ion channels in the post-synaptic membrane, allowing these ions (i.e. sodium - NA+, Ca++) to flow, carrying their positive charge, into the cell and triggering an EPSP.
Inhibitory transmitters (such as gamma-amino-butyric acid--GABA, serotonin--5-HT, or dopamine--DA) are those which affect specific channels, such as chloride and potassium channels in the post-synaptic membrane, allowing chloride to follow its concentration gradient (overcoming the opposing charge gradient) into the post-synaptic neuron and allowing potassium to follow its concentration gradient (overcoming its opposing charge gradient) and moving out. These ion movements make the inside of the neuron more negative, making an IPSP.
Notice that DA is listed as both excitatory and inhibitory. That's because some post-synaptic DA receptors are inhibitory, leading to IPSPs when activated by DA, and other DA receptors are excitatory, producing EPSPs when they bind with DA. In fact, many neurotransmitters have both excitatory and inhibitory receptors. The particular distribution of these two types of receptors on a postsynaptic cell will ultimately determine whether excitation or inhibition occurs when neurotransmitter travels across the synapse. Thus,we must be careful when referring to a particular transmitter as excitatory or inhibitory, because they probably can be both.
In summary,
- Excitatory postsynaptic potentials (EPSPs): a graded depolarizing (increasing) current that causes the membrane potential to become more positive and move closer to the threshold of excitation (approximately -55 millivolts) for action potential production.
- Inhibitory postsynaptic potentials (IPSPs): a hyperpolarizing (decreasing) current that causes the membrane potential to become more negative and move farther from the threshold of excitation (approximately -55 millivolts) for action potential production.
Spatial and Temporal Summation
There is one additional factor in this process. Each neuron connects with numerous other neurons, often receiving multiple impulses from them. Sometimes, a single excitatory postsynaptic potential (EPSP) is strong enough to induce an action potential in the postsynaptic neuron, but often multiple presynaptic inputs must create EPSPs around the same time for the postsynaptic neuron to be sufficiently depolarized to fire an action potential. Summation, either spatial or temporal, is the addition of these impulses at the axon hillock. Together, synaptic summation and the threshold for excitation act as a filter so that random “noise” in the system is not transmitted as important information.
At any moment in time, each neuron can be receiving mixed messages, both EPSPs and IPSPs. Thus, receiver neurons (post-synaptic neurons) can receive multiple inputs--simultaneously or over time or space. The multiple inputs can add up--this is called summation and there are two types. Spatial summation refers to two or more PSPs arriving at different locations (i.e. different spaces on the receiving neuron, thus spatial summation) on the post-synaptic (receiver) neuron simultaneously or close enough in time so that their voltages add together. For example, an IPSP of negative 25 millivolts (thousandths of a volt) might add to an EPSP of 50 millivolts occurring at the same time = 50 millivolts positive - 25 millivolts = net positive 25 millivolts. That would be spatial summation. The other kind of summation is temporal summation when the PSPs from a single pre-synaptic source, arriving to the post-synaptic neuron in "rapid fire," add together. For example, three EPSPs of 10 millivolts each occurring in rapid sequence would add together. Spatial and temporal summation are important in determining whether "trigger threshold" will be reached in the receiving neuron, triggering an action potential in that neuron. Summation permits activity from many input neurons to be integrated in the neuron receiving the inputs. If membrane depolarization does not reach the threshold level, an action potential will not happen. See Figure \(\PageIndex{3}\).
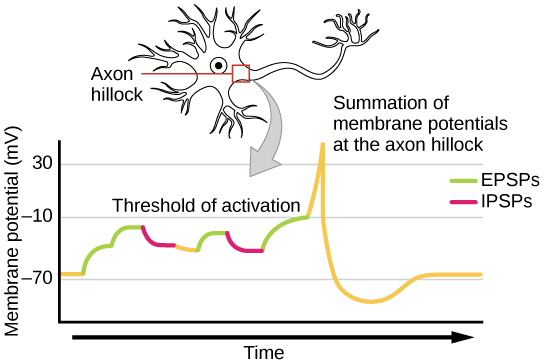
One neuron often has input from many presynaptic neurons, whether excitatory or inhibitory; therefore, inhibitory postsynaptic potentials (IPSPs) can cancel out EPSPs and vice versa. The net change in postsynaptic membrane voltage determines whether the postsynaptic cell has reached its threshold of excitation ("trigger threshold") needed to fire an action potential. If the neuron only receives excitatory impulses, it will also generate an action potential. However, if the neuron receives both inhibitory and excitatory inputs, the inhibition may cancel out the excitation and the nerve impulse will stop there. To review, spatial summation means that the effects of impulses received at different places on the neuron add up so that the neuron may fire when such impulses are received simultaneously, even if each input on its own would not be sufficient to cause firing. Temporal summation means that the effects of impulses received at the same place can add up if the impulses are received in close temporal succession. Thus, the neuron may fire when multiple inputs are received, even if each input on its own would not be sufficient to cause firing.
The Action Potential (the "nerve impulse")
The action potential is the main "output" message of the neuron and occurs in the axon where it travels from the beginning segment (axon hillock) to the end (axon terminal). The action potential is generated in a post-synaptic neuron only if a "trigger threshold" ("threshold of excitation") of approximately -55 millivolts is reached. Note that the trigger threshold varies from neuron to neuron and may be anywhere from minus 65 to minus 55, but will always be the same for any particular neuron. When that trigger voltage is attained (for example, as a result of an EPSP of at least 15 millivolts), then all the voltage-gated sodium ion (Na+) channels in the axon hillock suddenly open, allowing a massive inflow of Na+ into the axon along both concentration and electrical gradients for sodium. This produces a rapid, large positive shift or "spike" in the voltage of the post-synaptic neuron. This is the nerve impulse or action potential. In most neurons, it is a positive shift of about 100 to 130 millivolts, if we measure from the -70 millivolts of the resting potential, up to about a positive 30 to 60 millivolts, depending on the neuron. This value for any particular neuron is always the same for that neuron. Typically neurons with larger diameters produce the largest action potentials, while the neurons with smaller diameter axons produce action potentials in the lower range.
As fast as the voltage of the action potential rises, it starts to fall just as quickly after reaching its peak (corresponding to peak Na+ concentration inside the neuron). It quickly falls back to the resting potential and even a bit below (the so-called refractory period), as Na+ and K+ ions move out, before the return of the neuron's potential back to -70 mv, the resting potential. This rapid rise to the action potential's peak and then its rapid fall gives the action potential, when graphed, a spike appearance. For this reason, action potentials are often called "spikes" by neuroscientists. See the diagram of the action potential (Figure \(\PageIndex{4}\) & Figure \(\PageIndex{5}\)) to get a clearer picture of these events. Note that an EPSP, an excitatory post-synaptic potential, moves the neuron's voltage closer to "trigger threshold", increasing the chances that the neuron will be sufficiently "excited" to generate an action potential. By contrast, the IPSP, the inhibitory post-synaptic potential, moves the neuron's voltage further away from "trigger threshold" inhibiting the neuron from firing an action potential.
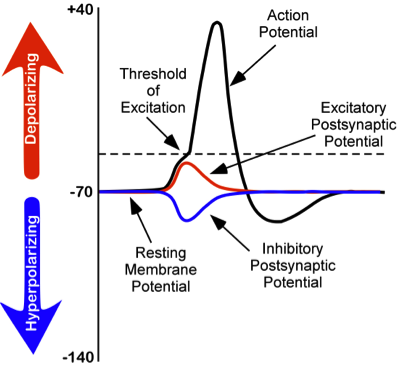
Figure \(\PageIndex{4}\): Changes in membrane potentials of neurons. (left) Dotted line represents trigger threshold ("threshold of excitation"), about -55 millivolts, an action potential is generated once trigger threshold voltage is reached.
Note that an EPSP (depolarization) moves the neuron's voltage more positive and thus closer to trigger threshold, making it more likely that the voltage reaches trigger threshold, "firing" an action potential down the axon of the neuron; thus it is excitatory. An IPSP does the opposite. It moves the voltage in the negative direction, further from trigger threshold, and thus inhibiting the neuron from producing an output (an action potential) in its axon. Also note that the peak of the action potential (top of the black line shaped like a spike) is the peak of its positive voltage and corresponds to the maximum concentration of Na+ ions inside the cell as a result of Na+ ion channels opening after trigger threshold has been reached. After this peak concentration of Na+ making the peak of the voltage of the action potential, positively charged ions (including K+ ions) begin to leave the interior of the cell and as they do so, the positive voltage inside the neuron progressively drops, corresponding to the downward slope of the spike. Note in addition that the spike goes further negative than the resting potential. In this state the neuron is inhibited by this refractory period and cannot fire another action potential for a brief time. This keeps the action potential as a discrete digital event. This is important for source coding, an important feature of neural coding, discussed in the chapter on learning and memory in this text. The entire process from the triggering of the action potential (which starts in the root of the axon, called the axon hillock, nearest the cell body) to the end of the refractory period takes about 1 millisecond, making the maximum rate at which a neuron can generate and "fire" action potentials (nerve "impulses") about 1,000 per second, although most neurons when active fire at a much lower frequency. Frequency of action potentials is one code that the nervous system uses to represent information. For example, the brighter a light source is, the higher the frequency of action potentials in the optic nerve in response to it.
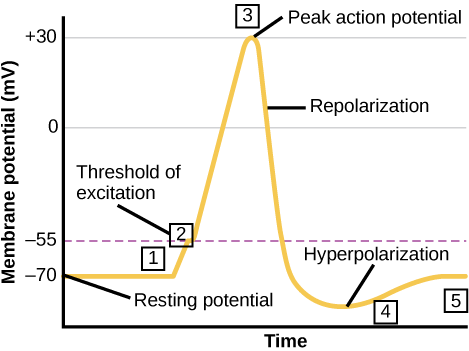
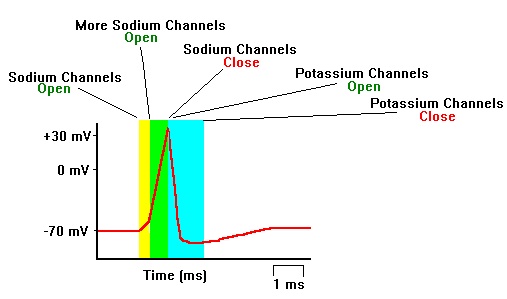
Figure \(\PageIndex{5}\): Formation of an action potential. (left) The formation of an action potential can be divided into five steps. (1) A stimulus from a sensory cell or another neuron causes the target cell to depolarize toward the threshold potential. (2) If the threshold of excitation is reached, all Na+ channels open and the membrane depolarizes. (3) At the peak action potential, K+ channels open and K+ begins to leave the cell. At the same time, Na+ channels close. (4) The membrane becomes hyperpolarized as K+ ions continue to leave the cell. The hyperpolarized membrane is in a refractory period and cannot fire. (5) The K+ channels close and the Na+/K+ transporter (requiring energy expenditure) restores the resting potential. (right) Sequence of opening and closing of Sodium (Na+) and Potassium (K+) ion channels producing the rising and falling phases of the action potential. (Image on left and caption from Lumen Boundless Biology; How Neurons Communicate; https://courses.lumenlearning.com/bo...s-communicate/. Unless otherwise noted, content is licensed under the Creative Commons Attribution 4.0 License. Image on right from Wikimedia Commons; File.مراحل ارسال سیگنال عصبی.jpg; https://commons.wikimedia.org/wiki/F...8%A8%DB%8C.jpg; by Vidakarimnia; licensed under the Creative Commons Attribution-Share Alike 3.0 Unported license).
Neural Conduction in Axons
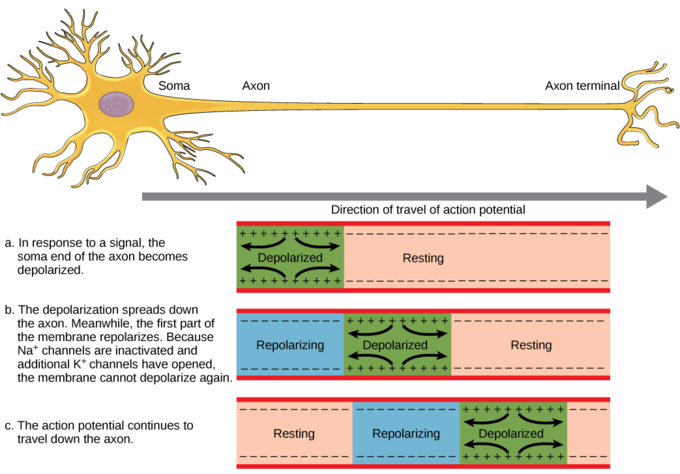
Once the trigger threshold ("threshold of excitation") is reached (dotted line in Figure \(\PageIndex{5}\)) and an action potential is generated, it is then conducted down the length of this neuron's axon (See Figure \(\PageIndex{6}\)). This conductance is due to the recreation of the full action potential at every sequential segment or patch of axon. This is caused by the opening of the voltage-dependent sodium and potassium channels located along the length of the axon. If the axon is myelinated it will travel more quickly than an unmyelinated one due to saltatory conduction (discussed below).
Once the AP reaches this neuron's axon terminal, its arrival causes the release of neurotransmitter molecules located there. Now this neuron is no longer called a post-synaptic neuron, but becomes a pre-synaptic neuron (a sender neuron) with respect to the next cell in line. It's this release of transmitter that leads to Between-Neuron communication, which is discussed in the next section of this chapter..
Saltatory Conduction
As discussed previously, many axons in the nervous system are coated in packets of the fatty substance, myelin which is produced by certain glial cells. This wrapping acts similar to insulation on an electrical wire and will speed up the process of action potential movement from the axon hillock to the terminals. These myelin wrappings do not fully ensheath the axon, but leave gaps. A node of Ranvier is one of these natural gaps in the myelin sheath along the axon. These unmyelinated spaces are about one micrometer long and contain voltage-gated (i.e., opened by voltage) sodium (Na+) and potassium (K+) ion channels (ions are electrically charged atoms). The flow of ions through these channels, particularly the Na+ channels, regenerates the action potential over and over again along the axon at each successive node of Ranvier. Below the myelinated sections, the positive charges move rapidly down the axon toward the next node to reproduce the action potential. This makes it appear that the action potential “jumps” from one node to the next, giving it the Latin name related to dancing, saltatory conduction.
Nodes of Ranvier also save energy for the neuron since the ion channels only need to be present and opened and closed at the nodes and not along the entire axon. It is extraordinary that the nodes are placed along the axon's length at just the right spatial intervals to make impulse conduction down the axon the most efficient and speedy as possible. One can only wonder at the incredible precision with which natural selection operated on this feature of myelinated axons over the long course of animal evolution to create this optimal spacing of the nodes.
Because the action potential in a myelinated axon must be regenerated fewer times to move any particular distance along the length of the axon, it reaches its destination faster, compared to the speed of conduction in an unmyelinated axon.
From Within-Neuron to Between-Neuron Communication and Beyond
A single neuron by itself can't generate psychological states like feelings, perceptions, or thoughts. Neurons must interact with other neurons. The generation of a thought, or other complex psychological/mental experience, requires that enormous numbers of neurons interact with one another. To interact, they must communicate with other neurons.
As described previously, neurons communicate with one another, across spaces between them known as "synaptic gaps" by the release of chemicals (called neurotransmitters or simply transmitters) from one neuron's axon terminals onto receptor sites of the target neuron (a post-synaptic neuron).
There are an enormous number of neurons in the human brain, but the number of possible different combinations of synaptic connections among those 80-100 billion neurons is unimaginable--one neuroscientist estimated that the number of possible patterns of interconnect between neurons in a human brain exceeds the number of atoms in the entire universe! This neural complexity appears to be sufficient to code all the information contained in a human brain including all the thoughts, feelings, perceptions and memories of a human lifetime. The foundational process of Between-Neuron (Synaptic) Communication is detailed in the next section.
Attributions
Communication within the Nervous System, "Neurons Generate Voltage Changes to Code Information" by Kenneth A. Koenigshofer, PhD, Chaffey College, is licensed under CC BY 4.0
Figure \(\PageIndex{4}\), and some text adapted from: Furtak, S. (2021). Neurons. In R. Biswas-Diener & E. Diener (Eds), Noba textbook series: Psychology. Champaign, IL: DEF publishers. Retrieved from http://noba.to/s678why4; Neurons by Sharon Furtak at NOBA is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License. Figure \(\PageIndex{4}\) caption by Kenneth A. Koenigshofer, PhD, Chaffey College.
"Key Points" and "Key Terms" adapted from: General Biology (Boundless), Chapter 35, The Nervous System;
https://bio.libretexts.org/Bookshelv...gy_(Boundless); LibreTexts content is licensed by CC BY-NC-SA 3.0. Legal.
Some text adapted from: General Biology (Boundless), Chapter 35, The Nervous System;
https://bio.libretexts.org/Bookshelv...gy_(Boundless); LibreTexts content is licensed by CC BY-NC-SA 3.0. Legal.
Neurons by Sharon Furtak, licensed under a CC BY-SA NC 4.0 International license.
Additional revisions made by Alan Keys, Ph.D., Sacramento City College, Sacramento, CA.