Although the theory of evolution by natural selection gained traction in scientific circles in the decades after Darwin’s publication of Origins, he was never able to discover the mechanisms that caused variation within members of the same species or the means by which traits were inherited. This began later in 1892 with the publication of The Germ-Plasm: A Theory of Heredity by August Weismann, the same Weismann of the mouse tail experiment presented earlier in this chapter. In his book, Weissman proposed a theory of germ-plasm, which was a precursor to the later discovery and understanding of DNA. Weismann specialized in cytology, a branch of biology devoted to understanding the function of plant and animal cells. Germ-plasm, he proposed, was a substance in the germ cells (what we would call gametes, or sex cells, today) that carried hereditary information. He predicted that an offspring inherits half of its germ-plasm from each of its parents, and claimed that other cells (e.g. somatic, or body, cells) could not transmit genetic information from parents to offspring. This thereby erased the possibility that acquired traits (which he argued resided in somatic cells) could be inherited (Zou 2015). This contribution to evolutionary theory was an important step toward understanding genetic inheritance, but a connection between genetics and evolution was still lacking.
A series of lectures by a deceased Augustinian monk named Gregor Mendel (1822–1884), originally published in 1865, changed that perspective (Moore 1993, 285). Although Darwin was unknown to Mendel, he began a series of experiments with pea plants shortly after the publication of Darwin’s Origins, aiming to add to evolutionary understandings of heredity. As Mendel bred different generations of pea plants that had differences in seed shape and color, pod shape and color, flower position, and stem length, he documented consistent expression of some variations over others in subsequent generations. He meticulously documented the statistics of each crossing of plants and the percentages of phenotypes that resulted, eventually discovering the concept of dominance and recessiveness of characteristics. He also documented that there is no blending of inherited characteristics. For example, pea pod colors in the offspring of two parent plants, one with yellow pods and one with green, were either yellow or green, not yellowish green. Mendel also discovered that characteristics are inherited and expressed independently of each other, meaning the color of the pea pod was not necessarily expressed in conjunction with the pod being wrinkled or smooth. The recognition of the importance of Mendel’s work began with its rediscovery by Hugo de Vries and Carl Correns, both of whom were working on hypotheses regarding heredity in plants and had arrived at conclusions similar to Mendel’s. Both published papers supporting Mendel’s conclusions in 1900 (Moore 1993). Research into the inheritance of characteristics continued through the next three decades, and by the close of the 1930s, no major scientific questions remained regarding the transmission of heredity through genes, although what genes did and what chemicals they were made of were still under investigation.
The Modern Synthesis refers to the merging of Mendelian genetics with Darwinian evolution that took place between 1930 and 1950. The basic principles of the synthetic theory were influenced by scientists working in many different fields, including genetics, zoology, biology, paleontology, botany, and statistics. Although there were differences of opinion among them, evolution came to be defined as changes in allele frequencies within populations. Genetic mutations, changes in the genetic code that are the original source of variation in every living thing, were believed to be random, the sources of phenotypic variation, and transmitted through sexual reproduction. These assertions were supported by a growing body of field and laboratory research, as well as new work in mathematics in the field of population genetics that defined evolution as numerical changes in gene frequencies within an interbreeding population from one generation to the next (Corning 2020). These changes in gene frequencies were argued to be the result of natural selection, mutation, migration (gene flow), and genetic drift, or random chance. Empirical research and mathematics demonstrated that very small selective forces acting over a relatively long time were able to generate substantial evolutionary change, including speciation (Plutynski 2009). Thus, the Modern Synthesis encompassed both microevolution, which refers to changes in gene frequencies between generations within a population, and macroevolution, longer-term changes in a population that can eventually result in speciation, wherein individuals from different populations are no longer able to successfully interbreed and produce viable offspring.
Genetics and the Origin of Species, published in 1937 by Theodosius Dobzhansky (Figure 2.13), was a cornerstone of the modern synthesis, applying genetics to the study of natural selection in wild populations, appealing to both geneticists and field biologists. Dobzhansky was interested in speciation, particularly in finding out what kept one species distinct from another and how speciation occurred. His research involved fruit flies, the species Drosophila pseudoobscura. At the time he began in the 1920s, biologists assumed all members of the same species had nearly identical genes. Dobzhansky traveled from Canada to Mexico capturing wild members of D.pseudoobscura, discovering that different populations had different combinations of alleles (forms of a gene) that distinguished them from other populations, even though they were all members of the same species. What, then, led to the creation of new species? Dobzhansky realized it was sexual selection. Members of the same species are more likely to live among their own kind and to recognize, and prefer, them as mates. Over time, as a result of random mutations, natural selection in a given environment, and genetic drift, meaning random changes in allele frequencies, members of the same population accumulate mutations distinct to their own gene pool, eventually becoming genetically distinct from other populations. What this means is that they have become a new species.
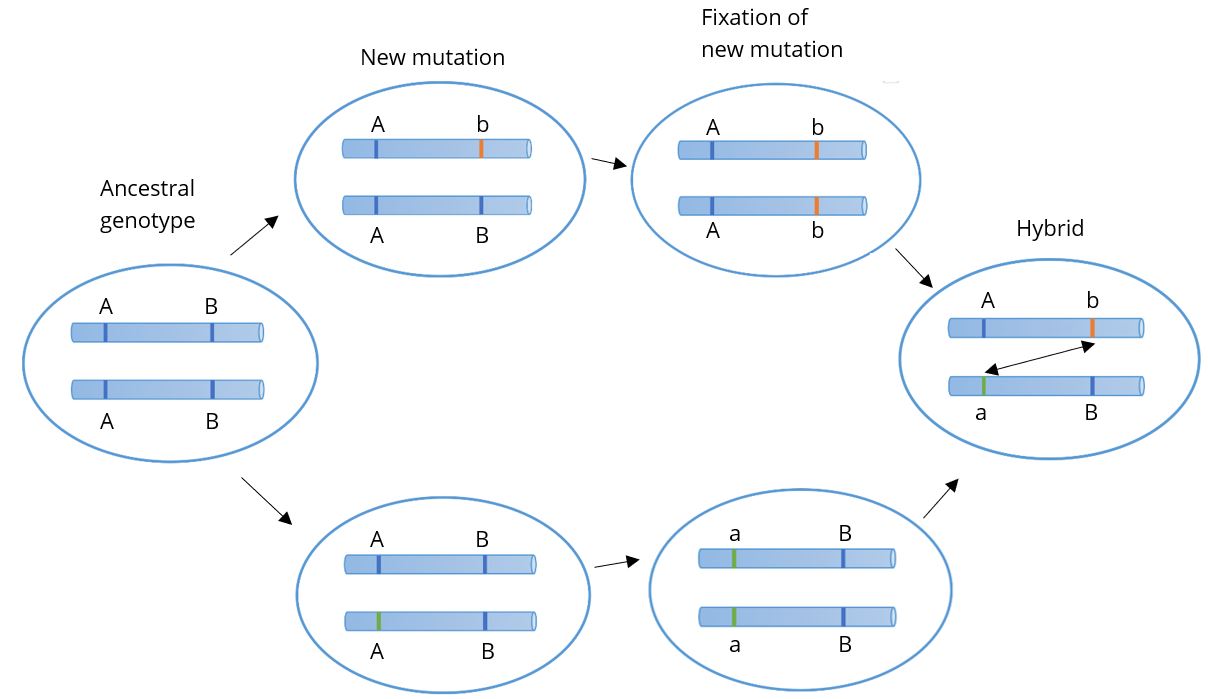
From these studies, Dobzhansky and others developed the Bateson-Dobzhansky-Muller model, also known as Dobzhansky-Muller model (Figure 2.14). It is a model of the evolution of genetic incompatibility. Combining genetics with natural selection, the model is important in understanding the role of reproductive isolation during speciation and the role of natural selection in bringing it about. Due to sexual selection (mate preference), populations can become reproductively isolated from one another. Eventually, novel mutations may arise and be selected for in one or both populations, rendering members of each genetically incompatible with the other, resulting in two distinct species.
Figure 2.14: The Dobzhansky-Muller Model: In the ancestral population the genotype is AABB. When two populations become isolated from each other, new mutations can arise. In one population uppercase A evolves into lowercase a, and in the other uppercase B evolves into lowercase b. When the two populations hybridize, it is the first time a and b interact with each other. When these alleles are incompatible, they represent Dobzhansky-Muller incompatibilities. Credit: Bateson-Dobzhansky-Muller model by OrientationEB is under a CC BY-SA 4.0 License.