The first successful sequencing of aDNA from an archaic hominin took place in 1997 with the sequencing of mitochondrial DNA (mtDNA) from a Neanderthal-type specimen from Feldhofer Cave. mtDNA is ideal for aDNA studies because it is more abundant than nuclear DNA in our cells. This mitochondrial sequence provided evidence that Neanderthals belonged in a clade separate from modern humans and that their mtDNA was four times more different from modern humans than modern human mtDNA was from each other (Krings et al. 1997).
Sequencing of nuclear DNA would not occur until more than ten years later. The first nuclear genomic sequence representing Neanderthals was produced by sequencing three individuals and using their sequences to create a composite draft Neanderthal genome (Green et al. 2010). The first high-coverage sequence of a single Neanderthal was that of a female Neanderthal who lived in Siberia, followed by another high-coverage sequence from a female Neanderthal whose remains were found in the Vidja cave in Croatia (Prüfer et al. 2014). High-coverage sequences are produced when the genome has been sequenced multiple times, which ensures that the sequences are a true reflection of the genomic sequence and not due to errors that occur during the process of sequencing.
Collecting and Sequencing aDNA
While aDNA can be collected from many different sources (e.g., soft tissue, hair, paleo feces, soils, and sediments), when studying ancient hominins it is most often collected from bone and teeth. Because extraction of aDNA requires destruction of part of the tissue, and the morphology of the skeletal element might be informative, care needs to be taken when deciding what is sampled. Multiple samples are often taken to allow repeat sequencing and demonstrate reproducibility of results. All samples must be minimally handled to avoid contamination.
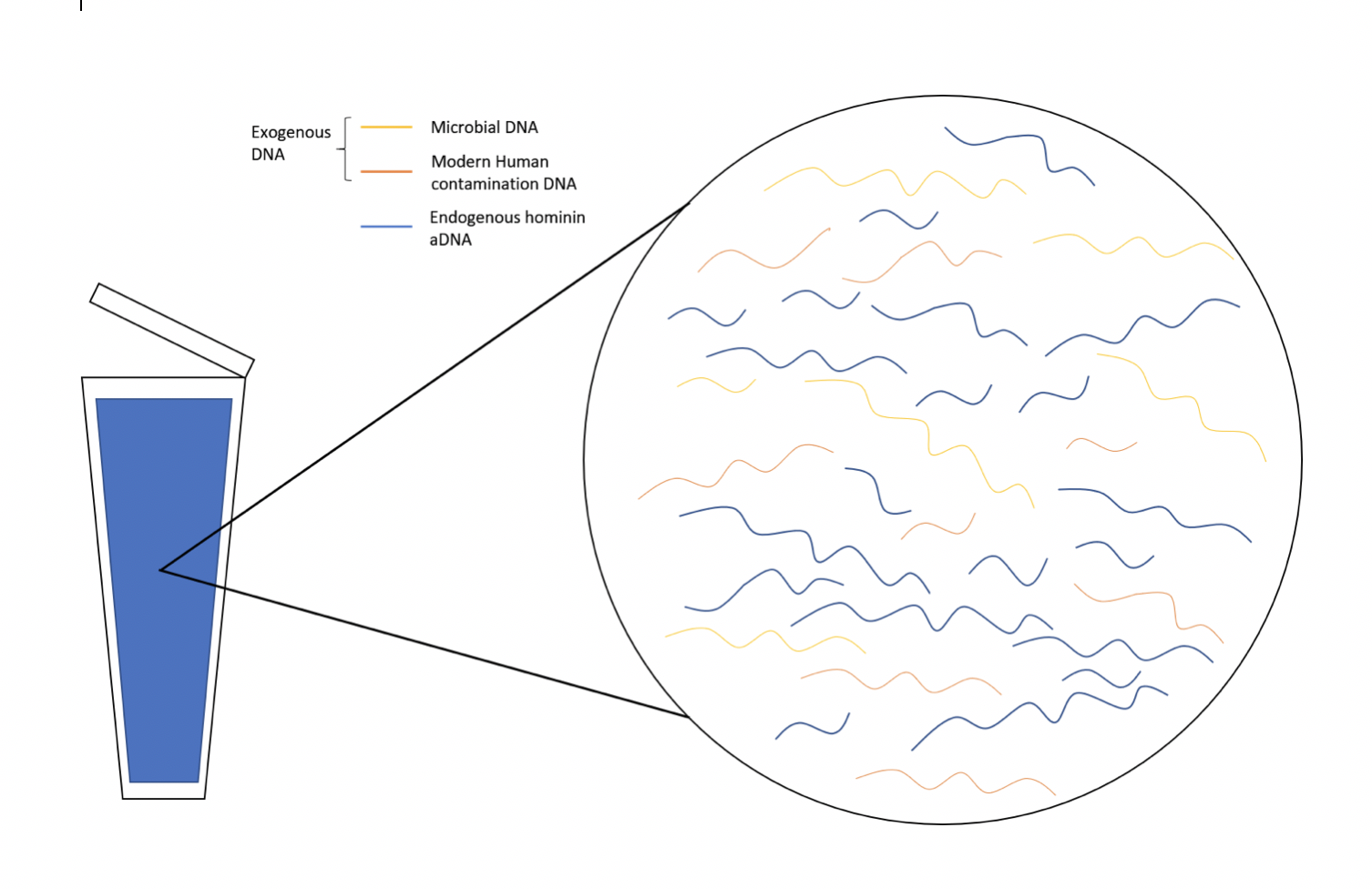
Endogenous aDNA, or DNA that was present in the tissue before the body decomposed, are usually in fragments 100 to 300 base pairs (bp) long due to degradation, and thus difficult to study. Sometimes DNA from other sources, known as exogenous DNA, are also found in samples. Some examples include DNA from microbes or modern human contamination (Figure D.1).
There are also modifications that occur to aDNA due to chemical reactions. For example, deamination results in Cytosine (C) to Thymine (T) conversions, which occur mostly at the 5’ end (5 prime end) of the DNA fragment. This in turn results in Guanine (G) to Adenine (A) substitutions on the 3’ end (3 prime end) of the DNA fragment. These sequence changes in aDNA might not reflect the original hominin sequence, yet these changes can be helpful when differentiating between aDNA and modern human DNA contamination. The environment plays a significant role, as DNA preserves well in cold conditions such as permafrost. aDNA has also been recovered from material found in drier environments under special conditions. Factors such as water percolation, salinity, pH, and microbial growth all affect the preservation of aDNA.
The bone that best preserves DNA after death is the petrous portion of the temporal bone. This forms part of the skull and protects the inner ear. Due to its high density, the petrous portion preserves DNA very well. Thus, it is possible to get DNA from older and less well-preserved individuals when using the petrous portion. Compared to other bones, the petrous portion not only preserves DNA better but also allows for the extraction of more DNA. The petrous portion can yield up to 100 times more DNA than other bones (Pinhasi et al. 2015)
Initially the short fragments and degraded nature of aDNA posed a big problem with the usual polymerase chain reaction (PCR) procedures used to sequence DNA. But, the advent of high-throughput sequencing has revolutionized sequencing the genomes of ancient hominins. High-throughput sequencing allows for the parallel sequencing of many fragments of DNA in one reaction, without prior knowledge of the target sequence. In this way, the maximum amount of available aDNA can be sequenced. Because the high-throughput sequencing method does not discriminate between endogenous aDNA and exogenous DNA, it is important to either ensure that there is as little contamination as possible and/or use methods that allow for differentiation of the target aDNA.