identify and describe functions of sleep-related brain structures
illustrate both directions of the shift between sleep and waking states
list the components of polysomnography (PSG)
describe rapid eye movement (REM) sleep and non-REM (NREM) sleep
determine sleep stages from PSG data
construct a diagram of healthy sleep architecture for eight hours of sleep
explain actigraphy, including its limitations compared to PSG
Introduction
Thanks in part to the availability of phone apps claiming to measure and analyze sleep, as well as an epic assortment of sleep analysis devices for the layperson to wear, we are experiencing a much-needed increase in the desire to deconstruct and explore our own sleep. To help understand the intricacies of sleep, let’s first get a fundamental understanding of the brain, at least in the context of how it functions when it’s awake versus sleeping. This will also be valuable in later chapters, which will make reference to assorted brain structures.
Brain Anatomy and Physiology
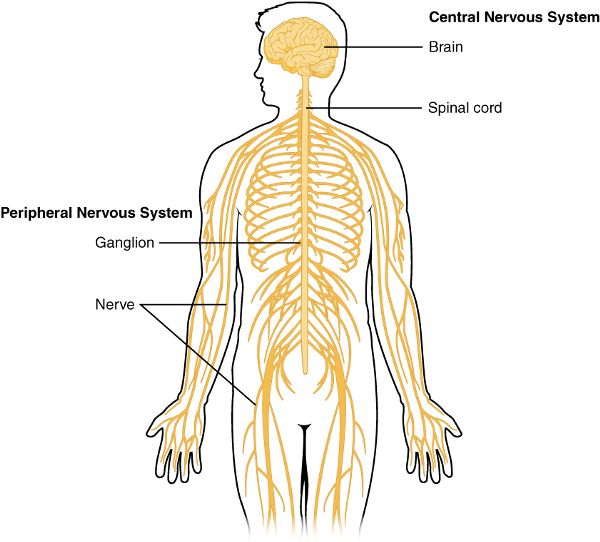
What molecules in your brain had to be released for you to make the decision to study this chapter? And how are you managing to hold your head up or read the words on the page? The nervous system carries signals through the body via neurons.1 These signals cause activity in muscles, glands, and other neurons. Some of the neurons are in the brain and the spinal cord, which together make up the central nervous system. Others travel throughout the rest of your body and comprise the peripheral nervous system (Figure 2.1). Sensory information from things we see, hear, feel, taste, or smell flows into the body and is processed by the central nervous system. After the brain has put us to sleep, it has a simple way of keeping most of that sensory information from awakening us. And while we are sleeping, the brain is actively creating the elaborate sleep architecture that carries us through the different stages and cascades necessary to secure the myriad benefits of a healthy night’s sleep.
Figure 2.1 Central and peripheral nervous systems
Note to reader: A more comprehensive review of brain anatomy and physiology is beyond the scope of this book, but this chapter will provide enough context and detail to give an understanding of sleep-related brain structures and functions. For additional brain anatomy and physiology, see The Brain from Top to Bottom, a website developed by Bruno Dubuc, hosted by McGill University in Canada, and labeled “copyleft” as a part of their desire to encourage people to freely copy and use their site’s content.2
The nervous system has two classes of cells: glial cells and neurons. Glial cells provide metabolic (metabolism = chemical reactions of the body) and physical support, while neurons carry the nervous system’s signals. Glial comes from the Greek for “glue.” Scientists chose this term when they noticed how numerous these cells were in the brain and mistakenly thought they had no purpose other than holding the neurons together. Later, it became clear that these cells are much more than brain glue and play a crucial role in preventing neurologic disorders through their sleep-related housekeeping activities. The misinformation surrounding glial cells did not end with their name. For ages, scientists believed glial cells immensely outnumbered neurons in the brain. Several studies suggested glial cells were ten times more numerous than neurons. However, in 2016, researchers from the Universidade Federal do Rio de Janeiro and University of Nevada School of Medicine used a new counting method and proposed that there are actually fewer glial cells than neurons in the brain. In their paper, they also provided a history of the techniques used to count glial cells, along with a discussion of the problems with the methods used that led scientists to the wrong conclusions for so many years.3 However, there are still some neuroscientists who debate this conclusion.
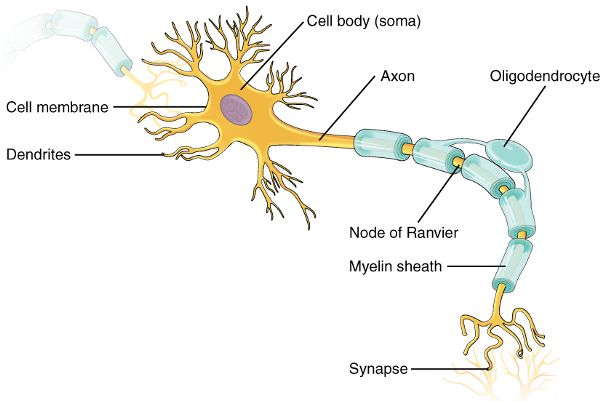
In contrast to glial cells, neurons use electrical activity and chemicals to carry signals throughout the body. The basic parts of a neuron are the dendrites, cell body, and axon (Figure 2.2). Dendrites carry information toward the cell body. From there, the signal travels to the axon to be transmitted to a muscle, gland, or another neuron. The functional connection between the neuron and the cell of its destination is called a synapse. Here, chemicals (neurotransmitters) or sometimes charged particles (ions) move from the first cell (presynaptic) to the second cell (postsynaptic). In this way, a signal, such as one triggered from the aroma of your roommate’s cooking, can make you aware of a delight to come. Meanwhile, another pathway, triggered by that same aroma, may cause you to salivate and activate your muscles to get you moving swiftly toward the kitchen so you can eat and fuel your brain for further studying.
Figure 2.2 Neuron and synapse
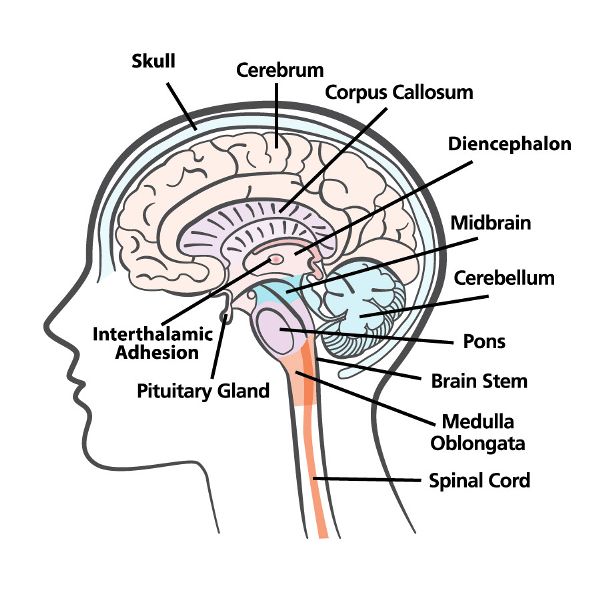
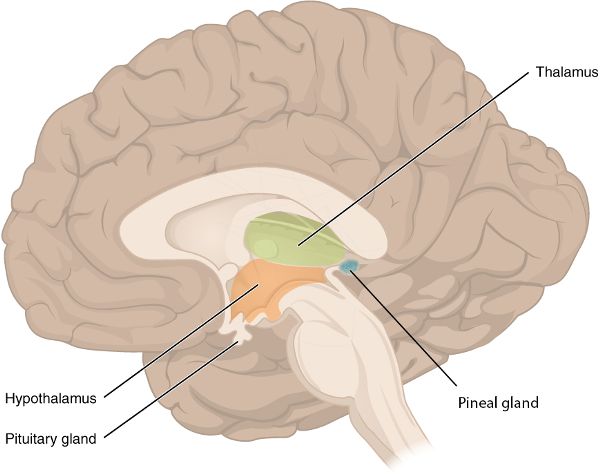
The four major parts of the brain are the brainstem, cerebellum, diencephalon, and cerebrum (Figure 2.3). The brainstem is continuous with and superior to (above) the spinal cord. Within the brainstem are the medulla oblongata, pons, and midbrain. Posterior to (behind) the brainstem is the cerebellum. The diencephalon—which includes the thalamus, hypothalamus, and epithalamus—sits on top of the brainstem. The cerebrum, the largest part of the brain, rests on top of the diencephalon.
Figure 2.3 The brain
Brainstem
With groups of neurons that control breathing, heart rate, and blood vessel diameter, the brainstem coordinates movements such as swallowing, coughing, sneezing, and much more. Pathways of sensory and motor information pass through and sometimes make connections in various regions of the brainstem. The reticular activating system (RAS)—a network of connections, primarily originating in the reticular formation—contains brainstem circuits that send signals to the cerebral cortex directly and also via the thalamus to contribute to consciousness (Figure 2.4). Sensory signals along this pathway keep you alert and oriented to your surroundings. The RAS is activated during awake states and is inactivated as part of initiating and maintaining sleep. However, when someone is sleeping, a strong enough sensory stimulus, such as a loud noise, will awaken the person via RAS activation. People differ from one another in the threshold required to activate the RAS during sleep: thus there are “heavy” and “light” sleepers. Signals from the eyes, the ears, and most of the rest of the body (e.g., temperature, touch, pain) travel through the RAS, but odors do not. This is why smoke detectors are important in sleeping areas. A person may die inhaling smoke from a fire while they are sleeping because the smell of smoke will not travel through the RAS and awaken them. If a person is unable to hear a fire alarm, they may consider smoke detectors that utilize extremely bright flashing lights or strong pillow vibrations to activate RAS pathways and increase their chances of awakening.
Figure 2.4 The brainstem
Cerebellum
Although the cerebellum is only one-tenth of the weight of the brain, it contains almost half of the brain’s neurons. Many of these neurons are dedicated to coordinating and optimizing movement, as well as maintaining posture and balance. While the preliminary motor signal to make a move, such as throwing a ball or saying a word, originates in the motor area of the cerebral cortex, that signal will loop into the cerebellum and back to the cerebral cortex. The benefit of the cerebellar input is that the movement will be smoother and more precise. There are also nonmotor functions of the cerebellum, such as learning and information processing, and a number of sleep-related functions. Research shows cerebral cortex and cerebellar interactions are crucial for memory consolidation, and some of these interactions occur particularly during sleep.4 Cerebellar activity also changes depending on the specific stage of sleep. Scientists continue to debate the exact role of the cerebellum in sleep, but it is clear that its dysfunction can cause sleep problems. In the presence of abnormal cerebellar function due to damage or a neurologic disorder, the sleep-wake cycle can be disrupted, and sleep disorders may be present. Of interest is that clock genes—regulators of the circadian rhythm—are expressed by cerebellar cells, but their function in this region remains to be elucidated.
Diencephalon
The thalamus—the largest part of the diencephalon—is a relay station, transmitting sensory information from the spinal cord and brainstem up into the sensory areas of the cerebral cortex (Figure 2.5). Additionally, by conveying information from the cerebellum and other brain structures up to the motor regions of the cerebral cortex, the thalamus is instrumental in creating coordinated movement. There are also thalamic functions associated with learning, memory, emotions, and consciousness. This consciousness is maintained in part by the thalamus transmitting some of the RAS signals up to the cerebral cortex. In contrast, during some components of sleep, the thalamus sends oscillatory signals to a large area of the cerebral cortex, in effect interfering with the cerebrocortical reception of sensory input that would normally travel up from the RAS. Oscillatory signals in this setting refer to neuronal electrical activity that is regular and synchronized, as opposed to neuronal sensory activity while awake, which would be irregular and not synchronized in a widespread manner.
Figure 2.5 The diencephalon shown in a mid-section view of the brain
Posterior and superior to the thalamus, the epithalamus contains the habenular nuclei, which associate emotions with smells—for example, the reaction you may have to the fragrance of your ipo (Hawaiian for “sweetheart”).
The other structure in the epithalamus is the pineal gland, a pea-sized structure that releases the hormone melatonin. Hormones are molecules that flow through the blood to their target structure, where they have an effect. This is the mechanism of action of the endocrine system.5 Therefore, the pineal gland, though it is in the brain, is a part of the endocrine system. During the darkness of night, the pineal gland releases its highest levels of melatonin, thereby regulating the circadian rhythm (see chapter 3).
Figure 2.6 Ipo
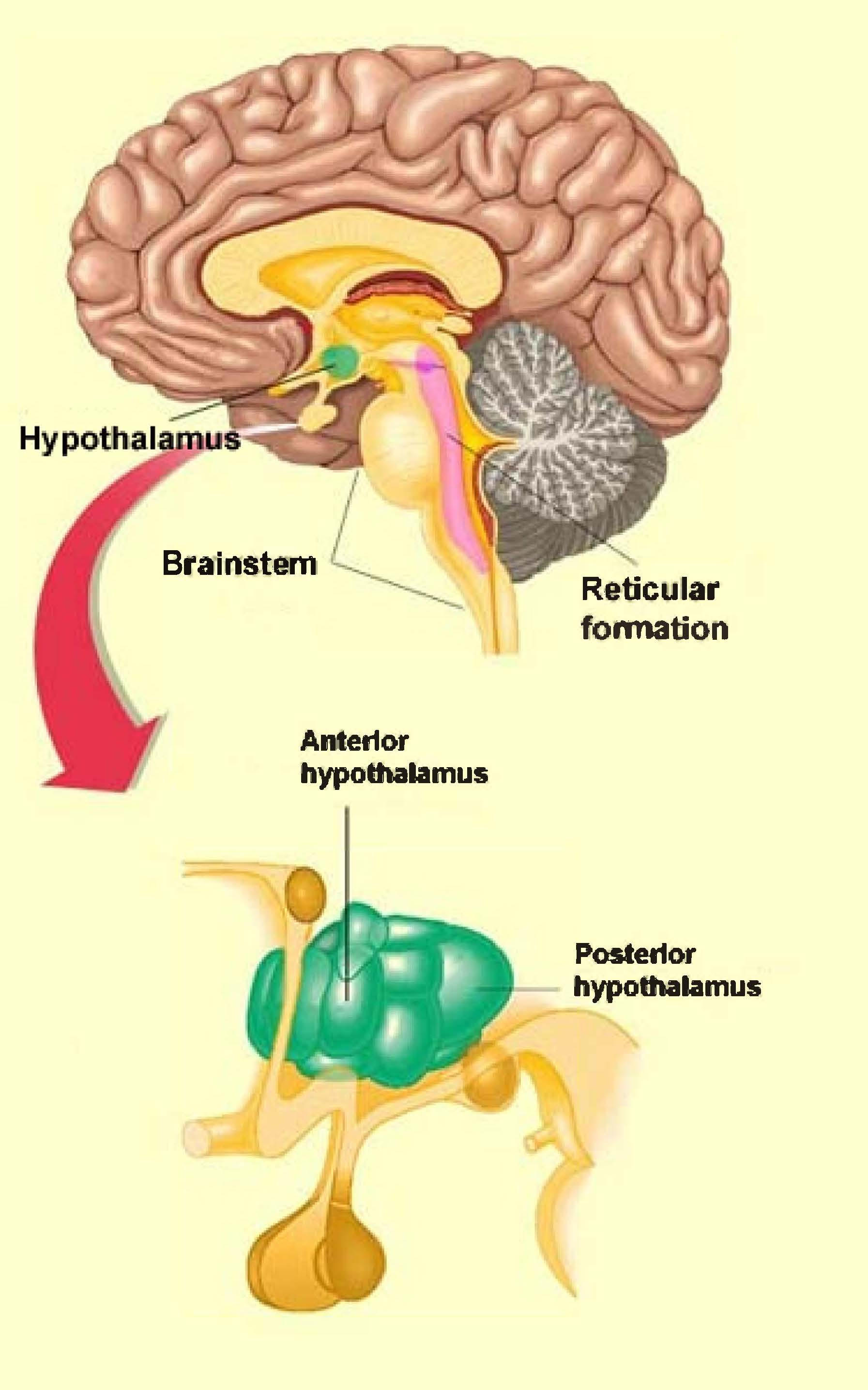
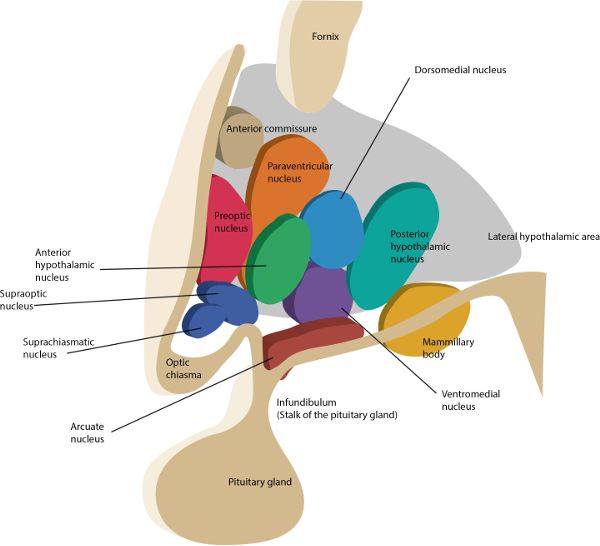
The hypothalamus (hypo = under) is made up of several nuclei with a vast array of functions. You may be familiar with the nucleus (plural, nuclei) as the part of a cell that contains the genetic material. However, in the brain, nucleus refers to a group of neuronal cell bodies such as those comprising the hypothalamus (Figure 2.7). To get a sense of the range of functions of the hypothalamus, they include—but are not limited to—regulating body temperature, generating the feeling of being satisfied after eating, being sexually aroused, changing heart rate, and controlling the circadian rhythm.
Of the many nuclei in the hypothalamus, the suprachiasmatic nucleus (SCN) is the one that orchestrates the circadian rhythm. As covered in chapter 3, the light- and dark-dependent signals from the eyes are one of the driving forces of the SCN, which regulates the pineal gland’s release of melatonin.
The posterior hypothalamus (posterior hypothalamic nucleus) is a nucleus that contributes to an elaborate network of structures involved with maintaining the awake state. One of the molecules the posterior hypothalamus releases to sustain wakefulness is histamine. This explains, in part, the drowsiness experienced when taking an antihistamine, found in many allergy medications, which blocks the effects of histamine. In fact, one of the wake-producing pathways of caffeine is associated with activating the release of histamine from these neurons. The posterior hypothalamus also releases gamma-aminobutyric acid (GABA) to maintain wakefulness. It does this by inhibiting neurons that would normally inhibit cerebral cortex activity. If you are thinking, “That sounds like a double negative,” you are correct. Think of it this way: The awake cerebral cortex is actively processing information, but that processing can be inhibited by neural pathways, thus resulting in sleep or drowsiness. But if those drowsiness-inducing pathways are inhibited by GABA from the posterior hypothalamus, then the brain will remain alert.
Figure 2.7 Nuclei of the hypothalamus
To understand one of the mechanisms for falling asleep, let’s consider what would happen if the posterior hypothalamus, and its wake-promoting effects, were inhibited. Since one of the posterior hypothalamus’ roles is to facilitate the transmission of information up to the cerebral cortex, then inhibiting the posterior hypothalamus would support sleep onset by reducing cerebrocortex information processing. The anterior hypothalamus (anterior hypothalamic nucleus) pulls this off via GABA. When the anterior hypothalamus is activated by the neurotransmitter serotonin, and if the timing is right in terms of circadian rhythm, the posterior hypothalamus is inhibited by the anterior hypothalamus, helping bring about the sleep state. The RAS is also inhibited from the anterior hypothalamus’ GABA activity, further reducing the likelihood that sensory information will have alerting effects on the cerebral cortex. Now the brain can fall asleep, mostly uninterrupted from the outside experience.
Cerebrum
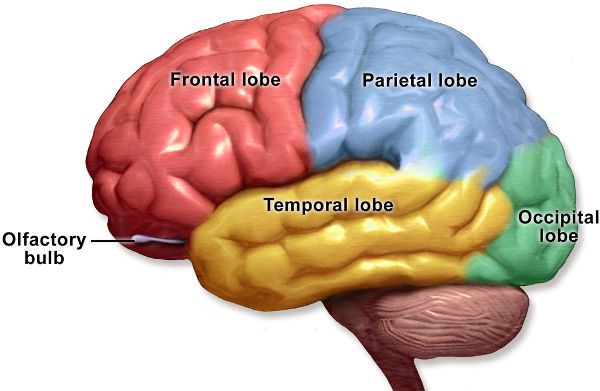
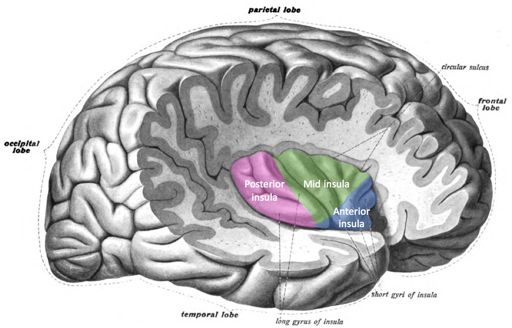
Singing a song, writing a story, playing a sport, and planning the day are made possible by our cerebrum. It is divided in half, with discrete regions that connect the left and right hemispheres. Deep inside the cerebrum are structures associated with an array of functions including memory, emotions, and motor control. The more superficial neurons of the cerebrum comprise the cerebral cortex, which is divided into four lobes: frontal, parietal, occipital, and temporal (Figure 2.8). The insula is another section of the cerebral cortex but is best visualized by creating a space between the meeting of the frontal and temporal lobes (Figure 2.9).
The frontal lobe contains areas for motor control, speech generation, odor identification, reasoning, personality, judgment, understanding consequences, learning complicated concepts, and more.
Figure 2.8 Lobes of the human brain
The parietal lobe receives sensory information, such as touch, temperature, pain, and itch. It also associates sensory data with other information, enabling you to identify a previously encountered item, such as your favorite fruit, entirely by touch. Part of the ability to understand language is also in the parietal lobe.
The occipital lobe processes visual information, including giving meaning to images. For example, image shapes coming from the eyes are combined in the occipital lobe in a manner that allows you to recognize your shoes solely by looking at them.
Figure 2.9 The insula
The temporal lobe receives and processes sounds and has areas for recognizing faces and perceiving smells.
The insula, previously one of the least understood brain regions, is now known to process taste, smell, sound, visceral and body surface sensations, and emotional responses such as empathy.
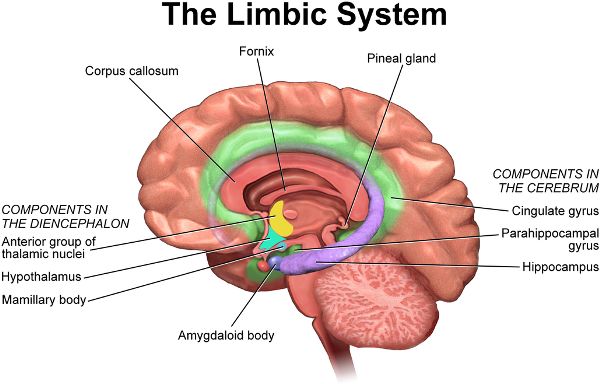
The limbic system includes part of the cerebral cortex and contains groups of neuronal cell bodies and pathways that interconnect cerebral cortex regions and other brain structures (Figure 2.10). It creates emotions such as pleasure, anger, and rage while also sparking drives for hunger and sex. The hippocampus, a vital structure for memory, is in the limbic system. The hippocampus has received more attention in recent decades because studies have suggested that the adult hippocampus produces new neurons, something previously deemed impossible anywhere in the adult brain. However, with further research, neuroscientists began questioning the existence of hippocampal neurogenesis. The debate has continued, with 2019 research swinging the view back in favor of neurogenesis in adult humans up to ninety years of age.6
Figure 2.10 The limbic system
In later chapters, we will revisit assorted aspects of brain anatomy, such as when learning about the creation and qualities of different types of dreams or how dreams can help us heal from trauma. For now, our discussion of brain activity will turn to how its characteristics are used to classify different waking and sleep states.
Figure 2.11 Polysomnogram
Polysomnogram
The polysomnogram (PSG) is the scientific tool for verifying sleep and is also used clinically to analyze sleep for disorders. While phone apps and actigraphy (see Actigraphy section) are commonly used to report sleep data of varying value, the scientific community has agreed to physiologically define sleep in humans as a set of stereotypical electrical signals from the brain, eyes, and skeletal muscles. Together, these three measurements—electroencephalogram, electrooculogram, and electromyogram—comprise the polysomnogram (poly = many, somno = sleep, gram = recording; Figure 2.11).
Electroencephalogram
During an electroencephalogram (EEG; electro = electricity, en = inside, cephalo = head, gram = recording), electrical activity in the brain travels through the skull and skin and can be detected by pasting tiny electrodes to the scalp (Figure 2.12). Viewing the voltage changes across time gives an indication of sleep onset and offset as well as the stage of sleep (such as REM or NREM, covered in the following section). The voltage change is measured vertically along the y axis, and the time change is measured horizontally along the x axis. This axis orientation is typical for all three types of polysomnogram recordings (electroencephalogram, electrooculogram, and electromyogram), but the scale on the y axis may vary.
Figure 2.12 Baby connected to EEG
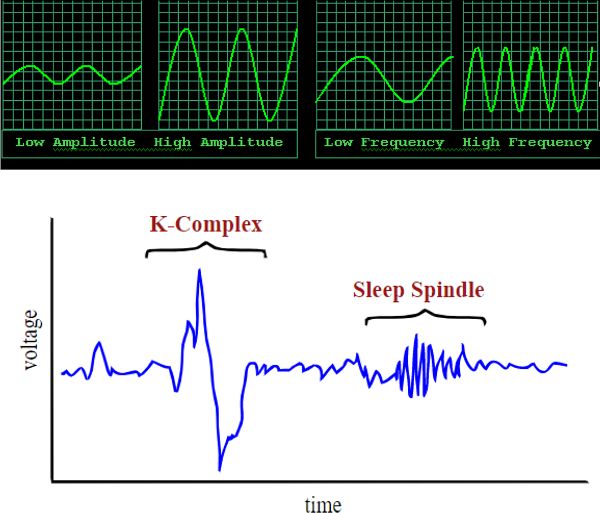
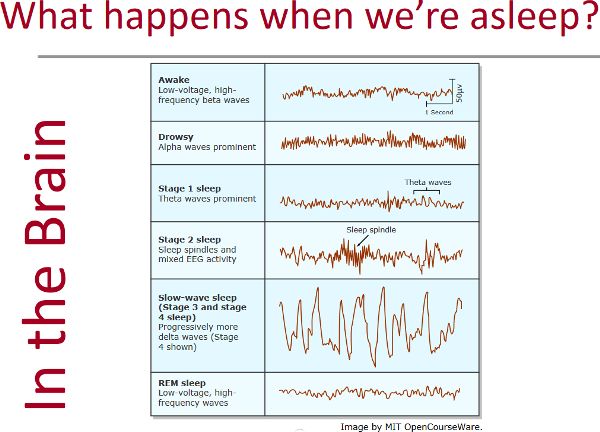
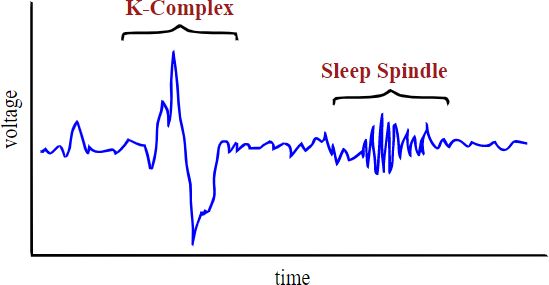
The PSG electrical wave characteristics are amplitude, frequency, and morphology (Figure 2.13). Wave amplitude is exactly what it sounds like: the size of the wave—a y axis measurement of voltage. Frequency describes how fast the waves are coming, so will be measured by looking along the x axis, at timing (Figure 2.14). The units for frequency are measured in hertz, also known as “cycles per second,” with a cycle being an entire wave. So this refers to how many whole waves are arriving every second. (The term hertz [Hz] was named after a person who studied electromagnetic waves.) Morphology (morph = form) is a way to look along the recording for unique shapes, such as a sleep spindle or K-complex, which are discussed in relation to NREM 2 in the Sleep Stages section (Figure 2.15). Different physiological states, such as sleeping or thinking, can be identified by EEG (Figure 2.16).
Figure 2.13 Wave amplitude and frequency as well as a K-complex and sleep spindleFigure 2.14 Waves in HawaiʻiFigure 2.15 Riding the perfect wave
Beta: awake, alert, thinking; 14–40 Hz
Alpha: awake, resting the mind, eyes closed; 8–13 Hz
Theta: drowsiness, daydreaming, sleep; 4–7 Hz
Delta: sleep; 1–4 Hz
Figure 2.16 EEG recordings
Electrooculogram
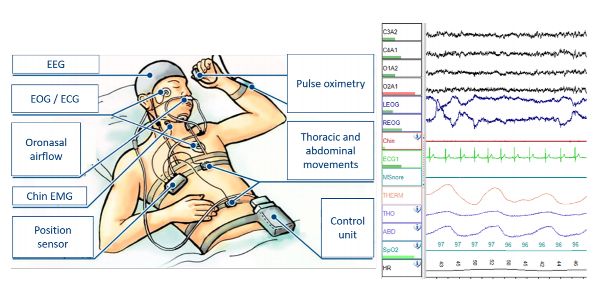
Different parts of the sleep cycle have particular eye movements that can be recorded by pasting electrodes on the skin beyond the outer corner of each eye for an electrooculogram (EOG; electro = electricity, oculo = eye, gram = recording). The anterior (front) region of the eyeball is positively charged compared to its posterior (back) region. This charge difference is utilized to generate a voltage trace for each eye, indicating if the eye is moved toward or away from the electrode, as well as the speed and size of the movements (Figure 2.17).
Figure 2.17 Polysomnography set-up and data
Electromyogram
Body movement during sleep can be categorized to determine sleep stages. Electrodes are typically placed below the chin and on the leg for an electromyogram (EMG; electro = electricity, myo = muscle, gram = recording). If you are sitting up reading this chapter and start to fall asleep, your head would fall slightly forward because the postural muscles below the chin relax. This change in muscle tone is picked up by an EMG. During a night’s sleep, it is normal to change position, twitch, and even have periods of paralysis. The EMG displays the type and timing of this movement (or lack of movement) so that data can be combined with the EOG and EEG to provide details about a person’s sleep.
Additional Clinical Measures
The EEG, EOG, and EMG are useful in research, but a clinical sleep study relies on additional physiological data. The sleep technician will connect the patient to devices to measure heart activity (electrocardiograph), blood oxygen (pulse oximeter), breathing effort (chest and abdominal expansion measurement instruments), and breath movement at the mouth and nose (oral/nasal airflow sensors). See the Apnea section of chapter 6 for a further discussion of these clinical measures.
Sleep Stages: REM Sleep and Non-REM Sleep
Sleep is divided into five major stages, each with an assortment of characteristics that distinguishes one stage from the other. However, they are named simply in reference to the presence or absence of rapid eye movement (REM). Curiously, REM sleep has only brief periods of rapid eye movement, but that name has persisted through the decades. Non-REM (NREM) sleep is further divided into four stages: NREM 1, 2, 3, and 4. Each of the five sleep stages occurs and repeats during different parts of a night’s sleep, comprising the full sleep cycle. The order, timing, and duration of the stages are referred to as sleep architecture. We will see that the brain has quite a job to do if it is to build a healthy night’s sleep according to the sleep architectural blueprint, which has been perfected over millennia.
REM Sleep
During REM sleep, we have vivid and emotional dreams while the body is paralyzed and not apparently regulating several physiological functions such as body temperature, heart rate, and blood pressure. REM sleep is composed of phasic and tonic components. Phasic REM sleep is easily recognized due to the “phases” when the eyes are darting back and forth. Tonic REM sleep, while still considered REM sleep, does not have eye movements but has similar brain activity to phasic REM sleep. Unless otherwise noted, in this textbook, REM will refer to REM in general without differentiating between phasic or tonic.
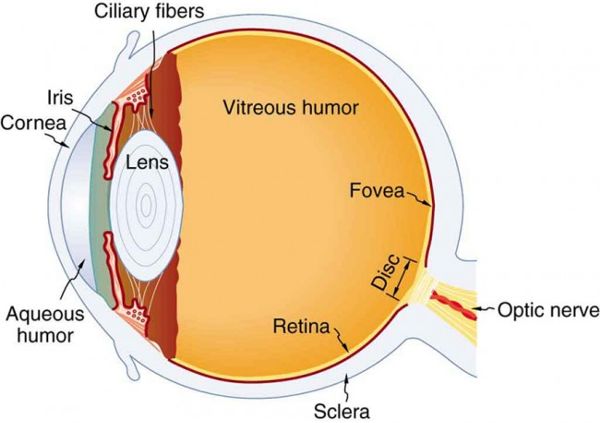
The purpose of those rapid eye movements may surprise you, especially if, like many others, you assumed the movements were associated with dream content (which they are not). Research by a Columbia University ocular physiologist suggests that rapid eye movement during sleep may be a way to keep the aqueous humor in the eyeball swirling in order to transport oxygen from the blood vessels of the iris to the cornea, which lacks blood vessels.8 During sleep, if the eyes did not move, the lack of aqueous fluid movement could result in corneal suffocation and cell death. When a person is awake, with the eyes open, there is a temperature difference on either side of the cornea that creates convection currents, causing the aqueous humor to move and transport the oxygen (Figure 2.18). The story gets more interesting when we try to understand why periods of REM sleep get longer throughout the night. The Columbia researcher’s group theorized that this lengthening of the REM sleep periods is necessary for oxygen transport, as the cumulative time (NREM + REM) the closed eye remains motionless increases from the first to the last hour of sleep.
Looking at the EEG of a person in REM sleep may lead you to believe they are awake because the electrical activity is asynchronous—it looks messy. This asynchronous activity is typical of the waking state, when the brain is processing myriad sensory input and thoughts.
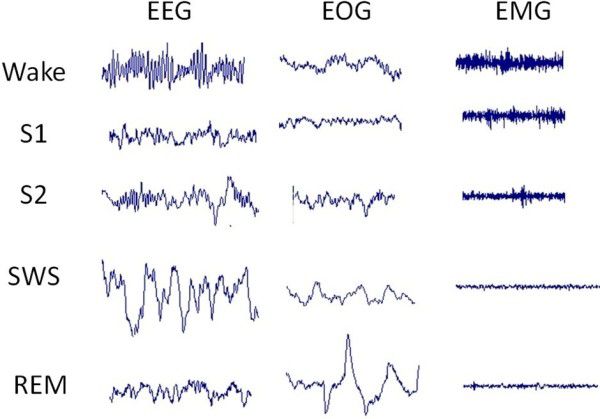
The flaccid paralysis of skeletal muscles during REM sleep leaves the person motionless except for breathing, rapid eye movement, and the occasional twitch, perhaps in a leg, finger, or facial muscle. There are also tiny skeletal muscles in the middle ear—providing protection from loud noises—that are not paralyzed during REM sleep, but this is certainly not observable to the casual viewer. Figure 2.19 shows the EEG, EOG, and EMG of the waking state and different stages of sleep: S1 (NREM 1), S2 (NREM 2), SWS (slow-wave sleep; NREM 3 and NREM 4), and REM.
Figure 2.18 The eyeFigure 2.19 EEG, EOG, and EMG
NREM Sleep
NREM 1 is how you enter sleep and is a light stage of sleep. Light sleep means a person is easily awakened. Many of us have been on one end of this experience: You wake up your friend, who is obviously sleeping, and you thoughtfully mention, “Sorry to wake you, but—” and they interrupt, “I was not sleeping!” and look at you like you said something ludicrous. If your friend happened to be hooked up to PSG, you would be able to show them they were in fact asleep. They may report they could not have been asleep because they were thinking about something, although usually something quite mundane. These “thoughts” are in fact the dull dreams of NREM 1. Another experience of NREM 1 can be when we lie down to sleep, and after a few moments, wonder why we were thinking something slightly absurd or illogical. We likely fell into NREM 1, easily awakened with no impression of being asleep, and then recalled the NREM 1 dream as a “thought.”
Here are some more facts about NREM 1 sleep:
• The EEG of NREM 1 is characterized by theta activity, with its lower frequency compared to the awake state.
• Although you may freak out your roommate by staring at them while they fall asleep (the sleep scientist’s folly), you can note when they drift into NREM 1 as their closed eyes have easily observed slow rolling movements.
• Occasionally, we see someone lying down, gently readjusting their position, and we conclude they are not sleeping. We may say something to them, to find they startle a bit and ask why we awakened them. These seemingly wakeful movements are normal in NREM 1 and are seen in the EMG. This stage can also include hypnic jerks, where the entire body or body parts have a large twitch, and there is often a sensation of falling. There is speculation that hypnic jerks are a vestigial response that prevented our ancestors who slept in trees from falling to the ground.
The term light sleep often refers to NREM 1 and NREM 2, but NREM 2 sleep, where you will spend almost half of your night, is more difficult to awaken from than in NREM 1. This is when things in the body start to slow down:
• The unique EEG morphology—sleep spindles and K-complexes—of NREM 2 makes it easy to differentiate this stage of sleep from the others (Figure 2.20). Sleep spindles may be associated with learning, and transferring information from short-to-long-term memory. K-complexes are generated in response to a stimulus, such as touch or sound, and may help us stay asleep during those potential disruptions.
Figure 2.20 NREM 2 EEG
• The eyes do not have any noticeable movements during NREM 2.
• There may still be some body movements during NREM 2, such as shifting position.
NREM 3 and NREM 4 together are often referred to as “deep sleep” because awakening from these stages is difficult and results in a fierce feeling of grogginess.
• The EEGs for NREM 3 and NREM 4 both contain large amplitude, slow waves—delta waves—giving both of these sleep stages the name “slow-wave sleep.” NREM 4 consists almost entirely of these slow waves, while NREM 3 has intermittent periods of the slow waves. Because this percentage of slow-wave sleep is the most noticeable difference between NREM 3 and NREM 4, many scientists have abandoned use of NREM 4, stating that NREM has only three stages, 1, 2, and 3. For simplicity and clarity in this text, we will use slow-wave sleep (SWS) to refer to NREM 3 and NREM 4 collectively, collapsing NREM 4 into NREM 3 when discussing the NREM stages.
• The eyes do not have any noticeable movements during SWS.
• Some body movement may occur during SWS, but it is minimal.
Some researchers debated the use of the word deep when referring to slow-wave sleep, so occasionally an article may seem contradictory to the convention. Which would you consider deep sleep: SWS, during which the body may be moving slightly and is still regulating many of its physiological functions, such as temperature and blood pressure, or REM sleep, when the body is paralyzed and not highly regulating some physiological functions, such as temperature and blood pressure? Ultimately, most have landed on considering SWS deep sleep due to the synchronous slow-wave brain activity and the difficulty of awakening a person from this stage, compared to the asynchronous brain activity of REM sleep and the relative ease of awakening from REM sleep.
Sleep Architecture
Sleep architecture is the timing and order of each of the sleep stages: REM and NREM 1, 2, and 3. Your brain and body are building something complex while you are lying there, and even something seemingly minor like a glass of wine shortly before bed is enough to disrupt your brain’s ability to create all the elements of sleep. Alcohol, a central nervous system depressant, is one of the many substances that can prevent the brain from generating some of the sleep stages, such as REM, and can wreak havoc on the body’s ability to organize the stages in a manner necessary to receive the benefits of a healthy night’s sleep.
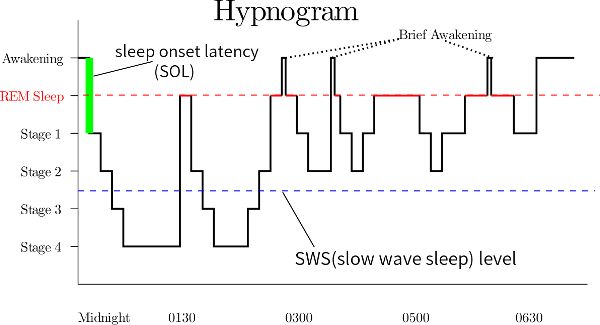
Sleep begins with NREM 1 and then moves through NREM 2 and 3 before going into the first period of REM, and this completes the first sleep cycle. On the way from NREM 3 to that first REM period, there may be some time in NREM 2 and 1. This first cycle takes about ninety minutes and will repeat throughout the night around five times, resulting in around 7.5 hours of sleep (consider doing that math to convince yourself it makes sense). The hypnogram in Figure 2.21 shows sleep architecture. Around midnight, this person took a few minutes to fall asleep (sleep-onset latency), went into NREM 1 (stage 1), and then went through each of the night’s sleep cycles before ultimately awakening fully at 6:30 a.m.
Figure 2.21 Hypnogram of sleep between midnight and 6:30 am
Within each ninety-minute cycle, as it repeats during the night, REM increases and NREM 3 and 4 decrease. Another way to think of this is that during the beginning of the night, you are getting more NREM 3 and 4, and during the last part of the night, you are getting more REM. Putting it all together, we also see that almost half the night is spent in NREM 2. (Note how the example hypnogram differentiates between NREM 3 and NREM 4 [as stages 3 and 4], while in this textbook, those two stages are typically merged into NREM 3.)
Actigraphy
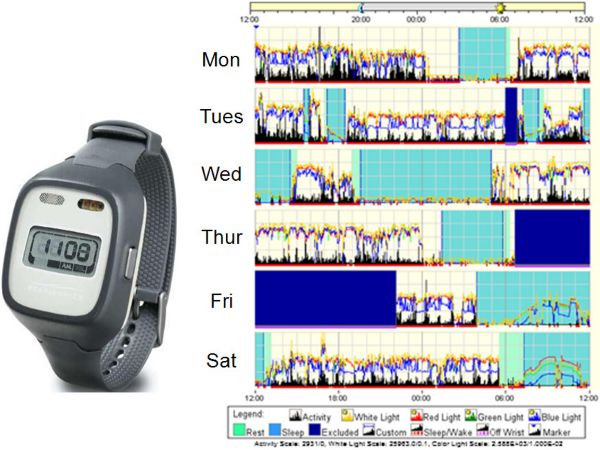
Cell phones contain a tiny instrument, an accelerometer, that changes the view on the phone display—the screen rotation—depending on how the phone is being held. In general, an accelerometer detects a change in the speed, direction, and size of a movement. Actigraphy utilizes accelerometers in small, watch-like devices to record a person’s physical activity, and consequently, in combination with computer algorithms, can be used to examine sleep in clinical and research studies (Figure 2.22).
The idea behind actigraphy is that during long enough periods of inactivity, a person must be sleeping, so that period would be labeled as sleep. Usually, the device will have a button that can be pressed when the person goes to bed and awakens. That context is helpful because sitting for two hours watching television could also seem a lot like sleep to an accelerometer. Polysomnography, with the three physiological measures of EEG, EOG, and EMG, has been used to validate actigraphy. However, it is important to understand actigraphy’s limitations. In actigraphy, we are using a device to measure movements and then making a leap utilizing computer programming to label different periods as sleep, while PSG is measuring the actual elements (EEG, EOG, and EMG) used in defining sleep.
Figure 2.22 Actigraphy device and data from a college student
The different measures and derivatives from actigraphy are as follows:
Sleep latency: how long it takes to fall asleep
Wake after sleep onset (WASO): how much time, after falling asleep, was spent awake
Total sleep time: from sleep onset to final awakening, with WASO subtracted
Sleep efficiency: total sleep time divided by the total time between sleep onset and final awakening; often referred to as sleep quality
Sleep latency should be at least fifteen minutes, as discussed in chapter 1, but certainly, much beyond that can begin to be frustrating. Sleep efficiency should be between 85–95 percent. To make this relatable, imagine that during the eight hours between falling asleep and waking up in the morning, you were awake for a few minutes enough times in the night that it added up to one hour of being awake (one hour of WASO). That would equate to seven hours of sleep during that eight-hour period. Dividing seven by eight gives a healthy sleep efficiency of 88 percent. Upon seeing their actigraphy data for the first time, many of my sleep science lab students are shocked by how many times they woke up during the night and even more surprised that it is considered normal and healthy. We are rarely aware of any of these awakenings.
What about a sleep efficiency of 100 percent—and why is that not included in the healthy range? With normal sleep architecture and a reasonable amount of sleep debt, a person would still occasionally awaken, as noted previously. However, if a person has a sleep disorder or an extreme amount of sleep debt, they may not awaken at all during their night’s sleep and have a sleep efficiency close to 100 percent.
3 Christopher S. von Bartheld, Jami Bahney, and Suzana Herculano-Houzel, “The Search for True Numbers of Neurons and Glial Cells in the Human Brain: A Review of 150 Years of Cell Counting,” Journal of Comparative Neurology 524, no. 18 (June 2016): 3865–95, doi.org/10.1002/cne.24040.
6 Elena P. Moreno-Jiménez et al., “Adult Hippocampal Neurogenesis Is Abundant in Neurologically Healthy Subjects and Drops Sharply in Patients with Alzheimer’s Disease,” Nature Medicine 25, no. 4 (March 2019): 554–60, https://doi.org/10.1038/s41591-019-0375-9.