
4.1: The Modern Synthesis and Population Genetics
- Page ID
- 177573
THE MODERN SYNTHESIS
How the first living organism came into being.
Genetically determined family lineages.
A family tree of all living organisms, based on genetic relationships.
The first living organism, from which all living things are descended.
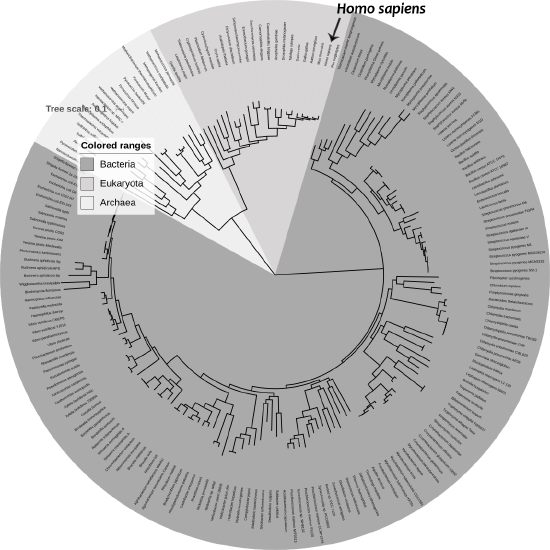 Figure \(\PageIndex{1}\): Phylogenetic tree of life.
Figure \(\PageIndex{1}\): Phylogenetic tree of life.Historical Framework
Rethinking Darwin
 Figure \(\PageIndex{2}\): The Modern Synthesis perspective: The ancestral population of giraffe had a range of variation in neck length. Those individuals with the longest necks would be the most likely to survive to pass on their longer-neck alleles to future generations.
Figure \(\PageIndex{2}\): The Modern Synthesis perspective: The ancestral population of giraffe had a range of variation in neck length. Those individuals with the longest necks would be the most likely to survive to pass on their longer-neck alleles to future generations. Figure 4\(\PageIndex{3}\): The Lamarckian hypothesis: If a short-necked parent giraffe often stretched its neck to reach higher branches, each generation of offspring would be born with somewhat longer necks.
Figure 4\(\PageIndex{3}\): The Lamarckian hypothesis: If a short-necked parent giraffe often stretched its neck to reach higher branches, each generation of offspring would be born with somewhat longer necks.An early model for inheritance that predicted that offspring inherit characteristics acquired during their parents’ lifetimes. This has now been proven incorrect.
An early model for inheritance that combines the Lamarckian idea of inheriting acquired characteristics with the idea that particles from different parts of the body make their way to the sex cells. This has now been proven to be incorrect.
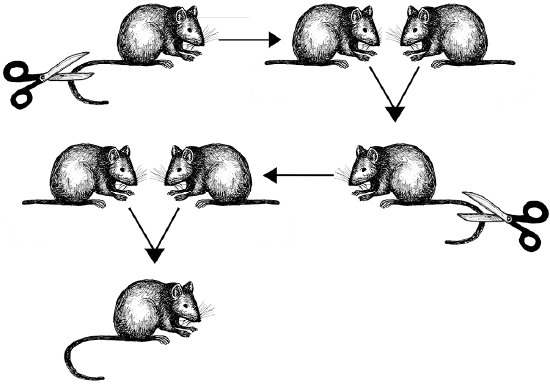 Figure \(\PageIndex{4}\): Weismann’s mouse-tail experiment showing that offspring do not inherit traits that the parents acquired during their lifetimes.
Figure \(\PageIndex{4}\): Weismann’s mouse-tail experiment showing that offspring do not inherit traits that the parents acquired during their lifetimes. Rediscovering Mendel
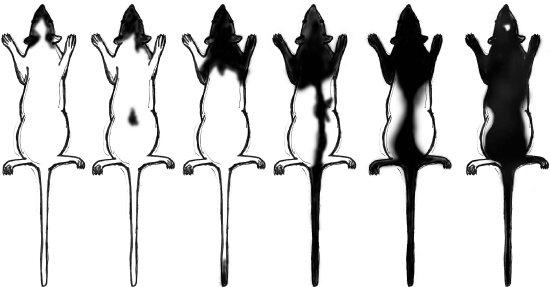 Figure \(\PageIndex{5}\): The continuous range of variation observed in hooded rat coat patterns after five generations.
Figure \(\PageIndex{5}\): The continuous range of variation observed in hooded rat coat patterns after five generations.A group of early biological scientists who believed that variation was caused by mutations in distinct, inherited cells.
A group of early biological scientists who believed that individual mutations of discrete hereditary units could never account for the continuous spectrum of variation seen in many traits.
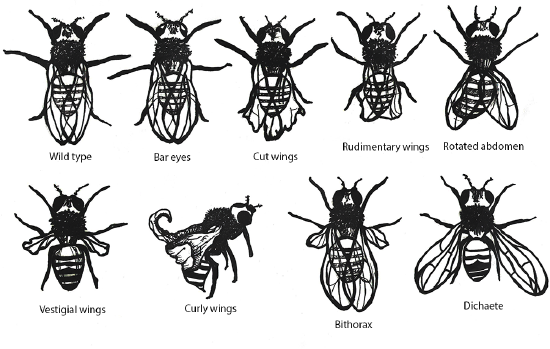 Figure \(\PageIndex{6}\): Examples of mutations producing phenotypic variation in a single species of fruit fly.
Figure \(\PageIndex{6}\): Examples of mutations producing phenotypic variation in a single species of fruit fly.Tying It All Together
Multiple forms of a trait; alternative phenotypes within a given species.
The integration of Darwin’s, Mendel’s, and subsequent research into a unified theory of evolution.
POPULATION GENETICS
Defining Species and Populations and the Variations Within Them
One of the major breakthroughs in understanding the mechanisms of evolutionary change came with the realization that evolution takes place at the level of populations, not within individuals. In the biological sciences, a population is defined as a group of individuals of the same species who are geographically near enough to one another that they can breed and produce new generations of individuals. Species are organisms whose individuals are capable of breeding because they are biologically and behaviorally compatible to produce viable, fertile offspring. Viable offspring are those offspring who are healthy enough to survive to adulthood. Fertile offspring can reproduce successfully to have offspring of their own. Both conditions must be met for individuals to be considered part of the same species. As you can imagine, these criteria complicate the identification of distinct species in fossilized remains of extinct populations. In those cases, we must examine how much phenotypic variation is typically found within a comparable modern-day species, and then determine whether the fossilized remains fall within the expected range of variation for a single species.
A group of individuals who are genetically similar enough and geographically near enough to one another that they can breed and produce new generations of individuals. A limited or restricted amount of gene flow between populations can occur due to geographical, cultural, linguistic, or environmental factors.
Organisms whose individuals are capable of breeding because they are biologically and behaviorally compatible to produce viable, fertile offspring.
Offspring that are healthy enough to survive to adulthood.
Offspring that can reproduce successfully to have offspring of their own.
Some species have subpopulations that are regionally distinct. These are classified as separate subspecies because they have their own unique phenotypes and are geographically isolated from one another, but if they do happen to encounter one another, they are still capable of successful interbreeding.
A distinct subtype of a species. Most often, this is a geographically isolated population with unique phenotypes; however, it remains biologically and behaviorally capable of interbreeding with other populations of the same species.
There are many examples of sterile hybrids that are offspring of parents from two different species. For example, horses and donkeys can breed and have offspring together. Depending on which species is the mother and which is the father, the offspring are either called mules, or hennies. Mules and hennies can live full life spans but are not able to have offspring of their own. Likewise, tigers and lions have been known to mate and have viable offspring. Again, depending on which species is the mother and which is the father, these offspring are called either ligers or tigons. Like mules and hennies, ligers and tigons are unable to reproduce. In each of these cases, the mismatched set of chromosomes that the offspring inherit still produce an adequate set of functioning genes for the hybrid offspring, but, once mixed and divided in meiosis, the gametes don’t contain the full complement of genes needed for survival in the third generation.
For the purpose of studying evolution, we recognize populations by their even smaller units: genes. Each individual, for genetic inheritance purposes, carries a collection of genes that can be passed down to future generations. For this reason, in population genetics, we think of populations as gene pools, which refers to the entire collection of genetic material in a breeding community that can be passed on from one generation to the next.
Remember, a gene is the basic unit of information that encodes the proteins needed to grow and function as a living organism. Each gene can have multiple alleles, or variants, each of which may produce a slightly different protein. For example, there are brown- or blue-pigment alleles for eye color (green is a slight variant of the brown type). The set of alleles that an individual inherits for a given gene is known as the genotype (e.g., inheriting both brown and blue eye pigments gives a genotype of Bb); while the observable traits that are produced by a genotype is known as the phenotype (e.g., a Bb individual exhibiting the dominant brown eye trait). For genes carried on our human chromosomes (our nuclear DNA), we inherit two copies of each, one from each parent. This means we may carry two of the same alleles (a homozygous genotype; eye pigment genotypes BB or bb) or two different alleles (a heterozygous genotype; eye pigment genotype Bb) for each nuclear gene. Only one of each of our alleles will get passed on to each of our children (the other will come from the child’s other parent). This means that children often inherit new genotypes and likely express unique phenotypes, compared to their parents. A common example is when two brown-eyed parents (who happen to be heterozygous for the pigment alleles) have a blue-eyed baby (genotype bb; who has inherited the recessive b alleles from both parents).
Defining Evolution
In order to understand evolution, it’s crucial to remember that evolution is always studied at the population level. Also, if a population were to stay exactly the same from one generation to the next, it would not be evolving. So evolution requires both a population of breeding individuals and some kind of a genetic change occurring within it. Thus, the simple definition of biological evolution is a change in the allele frequencies in a population over time. What do we mean by allele frequencies? Allele frequencies refer to the ratio, or percentage, of one allele (one variant of a gene) compared to the other alleles for that gene within the study population. By contrast, genotype frequencies are the ratios or percentages of the different homozygous and heterozygous genotypes in the population. Because we carry two alleles per genotype, the total count of alleles in a population will usually be exactly double the total count of genotypes in the same population (with the exception being rare cases in which an individual carries a different number of chromosomes than the typical two; e.g., Down syndrome results when a child carries three copies of Chromosome 21).
A change in the allele frequencies in a population over time.
The ratio, or percentage, of one allele compared to the other alleles for that gene within the study population.
The ratios or percentages of the different homozygous and heterozygous genotypes in the population.
REFERENCES
Castle, W. E., and J. C. Phillips. 1914. Piebald Rats and Selection: An Experimental Test of the Effectiveness of Selection and of the Theory of Gametic Purity in Mendelian Crosses. Carnegie Institute of Washington, No. 195. Washington, DC: Carnegie Institute of Washington.
Darwin, Charles. 1859. On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life. London: John Murray.
Dobzhansky, Theodosius (1937). Genetics and the Origin of Species. Columbia University Biological Series. New York: Columbia University Press.
Fisher, Ronald A. (1919). “The Correlation Between Relatives on the Supposition of Mendelian Inheritance.” Transactions of the Royal Society of Edinburgh 52 (2): 399–433.
Ford, E. B. 1942. Genetics for Medical Students. London: Methuen.
Ford, E. B. 1949. Mendelism and Evolution. London: Methuen.
Haldane, J. B. S. 1924. “A Mathematical Theory of Natural and Artificial Selection (Part 1).” Transactions of the Cambridge Philosophical Society 23:19–41.
Huxley, Julian. 1942. Evolution: The Modern Synthesis. London: Allen & Unwin.
Morgan, T. H. 1911. “Random Segregation Versus Coupling in Mendelian Inheritance.” Science 34 (873): 384.
Weismann, August. 1892. Das Keimplasma: Eine Theorie der Vererbung [The Germ Plasm: A Theory of Inheritance]. Jena: Fischer.
Wright, Sewall. 1932. “The Roles of Mutation, Inbreeding, Crossbreeding, and Selection in Evolution.” Proceedings of the Sixth International Congress on Genetics 1 (6): 356–366.
FIGURE ATTRIBUTIONS
Figure 4.1.1 Tree of life SVG by Ivica Letunic: Iletunic, retraced by Mariana Ruiz Villarreal: LadyofHats, has been designated to the public domain (CC0). This item has been modified (made grayscale, rotated, labels added).
Figure 4.1.2 Lamarckian Evolution original to Explorations: An Open Invitation to Biological Anthropology by Mary Nelson is under a CC BY-NC 4.0 License.
Figure 4.1.3 Modern Synthesis original to Explorations: An Open Invitation to Biological Anthropology by Mary Nelson is under a CC BY-NC 4.0 License.
Figure 4.1.4 Weismann’s mouse-tail experiment original to Explorations: An Open Invitation to Biological Anthropology by Mary Nelson is under a CC BY-NC 4.0 License.
Figure 4.1.5 Castle’s Hooded Rat Experiment original to Explorations: An Open Invitation to Biological Anthropology by Mary Nelson is under a CC BY-NC 4.0 License.
Figure 4.1.6 Morgan’s Mutant Fruit Flies original to Explorations: An Open Invitation to Biological Anthropology by Mary Nelson is under a CC BY-NC 4.0 License.